
Eligmodontia bolsonensis, Mares, Michael A., Braun, Janet K., Coyner, Brandi S., Van, Ronald A. & Bussche, Den, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181748 |
|
DOI |
https://doi.org/10.5281/zenodo.5621373 |
|
persistent identifier |
https://treatment.plazi.org/id/5F63A56A-FFD7-2F43-B9E0-FA8A3FFA4832 |
|
treatment provided by |
Plazi |
|
scientific name |
Eligmodontia bolsonensis |
| status |
sp. nov. |
Eligmodontia bolsonensis , new species
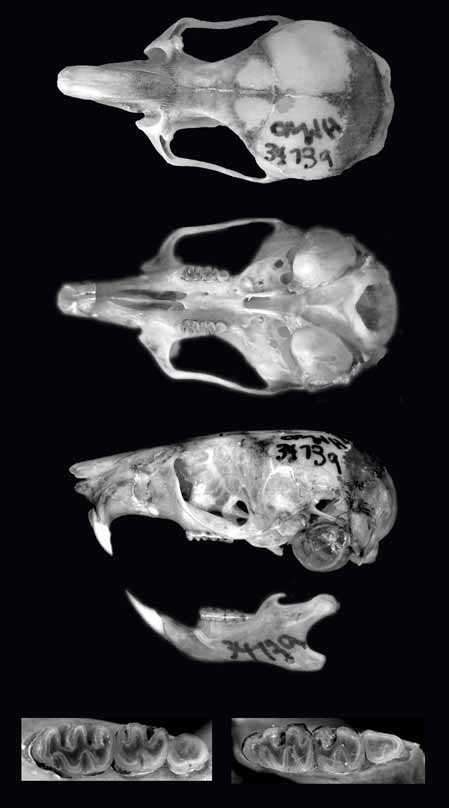
Holotype. OMNH 34739, adult female, collected by J. K. Braun on 8 October 1999 (original field number Arg 4924), skin, skull, skeleton.
Type locality. ARGENTINA: Catamarca Province: Pomán: Establecimiento Río Blanco, 28 km S, 13.3 km W Andalgalá 27°51’01”S, 66°18’17”W.
Distribution. This species is known presently from Catamarca Province, in the area north and west of the Sierra de Ambato and Sierra de Manchao from localities in the Salar de Pipanaco and Campo Arenal, and in the valley system extending north of Belẻn to Laguna Blanca. It also may occur in the valley system extending north-northeast through the Valles Calchaquíes to Cafayate, Salta Province ( Lanzone et al. 2007). This area of sand dunes and sand formations is relatively isolated from the Campo de Belẻn and the Salar de Pipanaco to the south by the Sierra de Belẻn.
Etymology. bolson—referring to a geographic feature + - ensis (L), adjectival suffix meaning “to belong to.” Named for the region in which it occurs—the Monte de Sierras y Bolsones.
Diagnosis. A member of the genus Eligmodontia distinguishable from all other species by the following combination of characters: dorsum light yellowish brown; venter immaculate; tail without pencil and whitish dorsally and ventrally; length of tail greater than length of head and body; darkish band extending from nose to between ears absent; size intermediate between E. moreni and E. typus (Table 5); greatest length of skull generally 23–25 mm; length of maxillary toothrow generally 3.7–4.0 mm; length of nasals generally 8.7–9.5 mm; anterior border of zygomatic plate generally slightly concave; bullae and eustachian tubes moderately developed; stapedial spines of bullae round or ovoid; knobs at frontoparietal suture well developed; braincase moderately inflated, but auditory bullae visible in dorsal view.
Measurement of holotype. External measurements (in mm): total length, 187; length of tail, 102; length of head and body, 85; length of hind foot, 23; length of ear, 16. Weight (in g): 17.5. Cranial measurements (in mm): greatest length of skull, 24.05; condylobasal length, 22.66; interorbital breadth, 3.69; zygomatic breadth, 12.21; breadth of braincase, 11.26; maxillary toothrow length, 3.68; bullar width, 4.81; bullar length, 4.32; palatilar length, 10.25; diastema length, 5.64; palatal bridge length, 4.18; palatal width at M1, 2.90; nasal length, 8.90; incisive foramen length, 5.23; incisive foramen width, 1.50. See Table 5 for measurements for all specimens examined.
Description. Size small as in other species in the genus. Pelage is long and lax. Overall coloration of the dorsum is very pale yellowish brown; hairs are light ochraceous buff for about the distal 1/2 and grayish for the proximal 1/2; long, dark guard hairs are present giving the dorsum a slightly darker appearance. Venter is immaculate and hairs are white to the base. Transition from dorsum to venter is sharply defined and delineated by a slightly brighter colored line of light ochraceous buff. Area around the nose and mouth is immaculate white. Ears have well-developed whitish pre-auricular tufts and moderately developed post-auricular patches. Fore- and hind feet are covered with whitish hairs; soles are lightly covered with hairs. Hind feet are elongated; hypothenar pad is absent; plantar pads D2-4 fused. Tail is unicolored, pale white above and below, longer than the length of the head and body, and without a pencil.
Skull delicate, rostrum slender, and braincase smooth and rounded ( Fig. 5 View FIGURE 5 ). Nasals not extending beyond premaxillary-frontal suture. Zygomatic arches delicate, parallel, and little expanded. Zygomatic notches moderately deep, greater than 1/2 the width of the zygomatic plate. Lacrimals well developed. Supraorbital edges divergent posteriorly, edges square, a distinct knob present at the frontoparietal suture. Frontoparietal suture crescent shaped. Interparietal well developed. Anterior border of zygomatic plate slightly concave. Masseteric tubercules at bases of zygomatic plates well developed. Palate long, extending beyond the posterior plane of M3. Incisive foramina long, the posterior margin about level with anterolabial and anterolingual conules of M1. Palatines with 1 or 2 small, round foramina. Posterolateral palatal pits anterior to mesopterygoid fossa. Bullae moderately inflated; eustachian tubes short, not reaching posterior edge of parapterygoid processes. Parapterygoid fossa shallow and broadly expanded laterally. Stapedial spine of auditory bullae rounded or ovoid. Alisphenoid strut present. Foramen ovale, carotid canal, stapedial foramen, and middle lacerate foramen present.
Posterior projection of the mandible with a notch (angular notch) not extending anterior to posterior edge of capsular projection. Knob of masseteric ridge exceeds dorsal edge of diastema of mandible. Caudal vertebrae number 30–31, the sacral vertebrae number 3, and the ribs number 13.
Upper incisors opisthodont, slender, ungrooved, and pigmented orange ( Fig. 5 View FIGURE 5 ). Maxillary toothrows slightly posteriorly convergent. Primary cusps of molars alternate. M1 with anteriomedian flexus obsolete or absent. Minor and major labial and lingual folds with lophs and styles absent or obsolete, including protostyle, parastyle, mesostyle, and enterostyle. M2 with hypoflexus and metaflexus open and lophs and styles absent; paraflexus present and well developed; posteroflexus absent; anteroloph well developed, extending greater than 2/3 the width of paracone and present even in well-worn teeth; paracone and metacone larger than protocone and hypocone. M3 with hypoflexus often present as a small notch; metaflexus distinct or present as an enamel island; anteroloph present, but may be obsolete in worn teeth; m1 with well-developed procingulum; anterolingual and anterolabial conulids not extending to lingual and labial edges of metaconid and protoconid; posterolophid and posteroflexid present and well developed, the former extending at least halfway to the lingual edge of entoconid; m2 with anterolabial cingulum, posterolophid, and posteroflexid present, except in worn teeth; m3 with hypoflexid present.
Comparisons. The only species of Eligmodontia currently known to be broadly sympatric with E. bolsonensis is E. moreni . We have collected the two species only a meter apart, with E. bolsonensis preferring soft, sandy substrates and E. moreni being found on the more dense clay and gravelly soils. In areas where E. bolsonensis does not occur, E. moreni is common on the soft, sandy soils. E. moreni may be distinguished by the following: larger size (Table 5); less buffy dorsum; posteriorly divergent band extending from the tip of the nose to between the ears, which appears more distinct due to the light coloration around the eyes; ears generally darker in coloration; bicolored, tufted tail; smaller knobs at the frontoparietal suture; larger bullae; eustachian tubes that extend anteriorly to the parapterygoid processes; narrower middle lacerate foramina; shorter, less developed stapedial spines of bullae flattened or adpressed to bullae; slightly shorter palate, which extends beyond the posterior border of M3 by the length of M3; and 2N=52, FN=50.
Eligmodontia typus occurs in the general region, but is not known to be sympatric with E. bolsonensis , although we have found several areas of sympatry between E. typus and E. moreni . E. typus may be distinguished by the following: smaller size (Table 5); darker dorsum; posteriorly divergent band extending from the tip of the nose to between the ears, which appears more distinct due to the light coloration around the eyes; general appearance of ears dark; bicolored tail without a tuft; knobs at the frontoparietal suture absent; shorter, wider nasals and rostrum; anterior border of zygomatic plate less concave; and braincase more inflated, the auditory bullae slightly visible in dorsal view.
TABLE 5. Measurements (mm) of species of Eligmodontia . Means, sample size, standard deviations, minimum and maximum are given for each measurement. Significant differences (t-test; P<0.05) are indicated as follows: a n. sp. vs. typus ; b n. sp. vs. moreni ; c typus vs. moreni .
continued.
Eligmodontia moreni Natural history. Reproductively active individuals were captured in October and November. Females were pregnant with 4, 5, and 7 embryos. Males had scrotal testes from 7 to 11 mm in length. Reproductively inactive males were collected in March and September.
Small mammal species captured at several localities with E. bolsonensis include Thylamys pallidior , Tadarida brasiliensis , Abrothrix andinus , Akodon sp., Andalgalomys olrogi , Calomys sp., E. moreni , E. puerulus , Graomys griseoflavus , Phyllotis xanthopygus , Ctenomys sp., Pipanacoctomys aureus , Microcavia australis , and Chaetophractus vellerosus .
Remarks. The karyotype reported by Lanzone et al. (2007) of 2N=44, FN=44 containing 1 pair of large metacentric chromosomes, 20 pairs of acrocentric chromosomes, a small submetacentric Y, and an acrocentric X, likely corresponds to this species. This species was listed as Eligmodontia typus marica by Massoia (1976/ 77), as Eligmodontia sp. 1 by Mares et al. (1997), and Eligmodontia marica by Lanzone et al. (2007).
Paratypes. Fourteen specimens collected at or near the type locality; 8 deposited in the OMNH (OMNH 34735, OMNH 34736, OMNH 34737, OMNH 34738, OMNH 34811, OMNH 34812, OMNH 34813, OMNH 34814) and 6 deposited in the CML (field numbers Arg 4914, Arg 4918, Arg 4923, Arg 4926, Arg 4928, Arg 4929).
Specimens examined (19). See Appendix 1.
Other referred specimens. ARGENTINA: Catamarca: Antofagasta de la Sierra: Pasto Ventura ( OMNH 34726/Arg 5200); 1.5 km S El Peñỏn ( OMNH 34725/Arg 5319). Belẻn: Agua de Dionisio, Hualfín 1 ( CML 872); 17 km N Barranca Larga OMNH 34727/Arg 4954, OMNH 34728/Arg 4964); 5.2 km S El Bolsỏn ( OMNH 34729/Arg 5168, OMNH 34730/Arg 5169); Laguna Blanca ( OMNH 34731/Arg 5025, OMNH 34732/Arg 5026); 1.7 km S Laguna Blanca ( OMNH 34734/Arg 5175, OMNH 34733/Arg 5176); La Salamanca, Hualfín ( Massoia 1976/77); Los Baños Termales, Hualfín, 3 ( CML 868, 869, 881); Pozo Jovita, Hualfín, 1 ( CML 875; Massoia 1976 /77). Santa María: 21 km SW El Desmonte ( OMNH 34743/Arg 4501); 33 km SW, 1 km N El Desmonte, 1 ( OMNH 22303); 7.9 km W jct. Hwy 47 and 40 ( OMNH 34806/Arg 4931, OMNH 34742/Arg 4932, CML /Arg 4933, CML /Arg 4934, CML /Arg 4935, CML /Arg 4936, OMNH 34807/ Arg 4937, OMNH 34808/Arg 4938, OMNH 34809/Arg 4939, OMNH 34810/Arg 4940, CML /Arg 4941, CML /Arg 4942, CML /Arg 4943, CML /Arg 4944); 9.9 km W jct. Hwy 47 and 40 ( OMNH 34740/Arg 4946, OMNH 34741/Arg 4947, CML /Arg 4949).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.