
Mantidactylus schulzi, Vences & Hildenbrand & Warmuth & Andreone & Glaw, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4486.4.10 |
|
publication LSID |
lsid:zoobank.org:pub:B3451229-F9F1-4C2A-B633-BC63DA79A727 |
|
DOI |
https://doi.org/10.5281/zenodo.5967783 |
|
persistent identifier |
https://treatment.plazi.org/id/5E3287C1-221E-FF8C-53AB-1295FE88F821 |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus schulzi |
| status |
sp. nov. |
Mantidactylus schulzi View in CoL sp. nov.
Figs. 3–4 View FIGURE 3 View FIGURE 4
Remarks. This species has previously been named Mantidactylus sp. aff. biporus "Tsaratanana Andampy" by Glaw & Vences (2007), Mantidactylus sp. aff. betsileanus by Andreone et al. (2009), Mantidactylus sp. 32 by Vieites et al. (2009) and Mantidactylus sp. Ca32 by Perl et al. (2014). The ICZN-LSID number for this nomenclatural act (description of a new species) is urn:lsid:zoobank.org:act:2739C501-C964-4FB2-9ADC- 0FE4CB85B05A.
Holotype. ZSM 653/2001 (field number FGMV 2001.150), adult male, collected on the Tsaratanana Massif, Manarikoba Forest, Andampy, "Camp 0" ( 14°02’32’’S, 48°45’42’’E, ca. 730 m above sea level), former Antsiranana province, northern Madagascar on 10–11 February 2001 by F. Andreone, F. Mattioli, J. Randrianirina & M. Vences.
Paratypes. ZMA 19374 (fgmv 2002.754, Adult female) and ZMA 19375 ( FGMV 2002.762 , Adult male), ZSM 821–822 View Materials /2003 (adult males) and ZSM 823–826 View Materials /2003 (adult females), all collected on 2 February 2003 at Manongarivo, Camp 0 ( 13°58’32’’S, 48°25’36’’E, 688 m above sea level), former Antsiranana province, northwestern Madagascar, by F. Glaw, R.D. Randrianiaina & M. Vences GoogleMaps ; ZSM 651–652/2001 and 654/2001, with same collection data as holotype. The field numbers of the zsm paratypes are provided in Table 1. Additional uncatalogued paratypes from the UADBA collection are FGMV 2002.749, FGMV 2002.756, FGMV 2002.758, FGMV 2002.760, FGMV 2002.761, FGMV 2002.763, FGMV 2002.764, and FGMV 2002.765, all with same collection data as ZMA 19374. These eight UADBA paratypes and the two ZMA paratypes were not available for morphological studies but had identical sequences as the holotype ( Fig. 1 View FIGURE 1 ).
Diagnosis. The new species is assigned to the genus Mantidactylus based on the presence of an intercalary element between terminal and subterminal phalanges of fingers and toes (verified by external observation), and of a central depression in femoral glands, presence of a rudimentary femoral gland in the female, and tympanum diameter distinctly larger in males compared to females. Within Mantidactylus , it is assigned to the subgenus Brygoomantis by the combination of (1) rather small body size (male SVL 21–24 mm, female SVL 25–29 mm), (2) absence of dorsolateral colour border, (3) absence of a distinct frenal stripe, (4) absence of large yellowish patches or stripes in the inguinal region or between colouration of flanks and belly, (5) only partially webbed feet, (6) riparian habits, living close to the water in streams, and (7) molecular phylogenetic relationships (see Glaw & Vences 2006).
In comparison with nine out of the 12 nominal species of Brygoomantis , according to data summarized from Glaw & Vences (2007) and Blommers-Schlösser & Blanc (1991), the new species is distinguished by its smaller body size (male SVL 21–24 mm): from M. bellyi ( 32–41 mm), M. bourgati (> 30 mm mm), M. biporus ( 26–27 mm), M. curtus (> 30 mm), M. ambohimitombi (> 40 mm), M. madecassus ( 27–30 mm), M. noralottae ( 33–35 mm), M. pauliani ( 25–32 mm), and M. ulcerosus ( 32–34 mm). Where advertisement calls are known, the new species also differs by advertisement calls: M. bellyi , M. biporus , and M. ulcerosus all emit pulsed notes with evenly spaced pulses (unevenly spaced in the new species) and as far as known with faster pulse rates, but detailed analyses and comparisons of their calls are still pending.
Two further species of Brygoomantis also are slightly larger, but with overlap of values: M. betsileanus (male SVL 23–28 mm), M. alutus ( 24–26 mm); M. schulzi sp. nov. is distinguished from M. alutus by a more distinct throat pattern, with a white central stripe on dark throat (vs. mostly light-coloured throat with indistinct dark mottling), by a weakly expressed light spot on the snout tip (vs. absence), and different advertisement calls (irregular short series of pulsed notes with a typical rate of 62–84 pulses per second, vs. a regular series of 5–14 pulsed notes of 19–22 pulses repeated at a pulse rate of about 50 per second); from M. betsileanus as defined by Blommers-Schlösser (1979) and Glaw & Vences (1994, 2007) by a less distinct white spot on the snout tip and much larger and more distinct femoral glands in males.
Finally, the new species is at first glance morphologically similar to M. tricinctus but strongly differs from this species by more extended webbing on the foot (almost absent in that species), and by a slightly larger body size (male SVL 21–24 mm, vs. 17–19 mm in M. tricinctus ). The calls of unambiguously identified M. tricinctus (from near the type locality in south-eastern Madagascar) remain undescribed and a bioacoustic comparison to the new species is therefore not possible.
Furthermore, the new species is distinguished from all nominal species of Brygoomantis (except M. ambohimitombi for which no genetic data are available) by strong genetic differences (pairwise 16S differences>6.7%).
Description of the male holotype. ZSM 653/2001 (field number FGMV 2001.150), adult male in mediocre state of preservation (softly fixed, similar to all other available specimens, Fig. 3 View FIGURE 3 ). For measurements, see Table 1. Body rather stout. Head wider than long, as wide as body. Snout rather pointed. Nostrils directed laterally, slightly protuberant, nearer to tip of snout than to eye. Canthus rostralis moderately distinct, loreal region slightly concave. Tympanum distinct, large, oval, horizontal diameter of tympanum 94% of horizontal eye diameter. Supratympanic fold distinct, beginning straight above, with rather distinct bend midway towards insertion of forelimb. Tongue ovoid, distinctly bifid posteriorly. Vomerine teeth form two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticular tubercles single. Outer metacarpal tubercle poorly recognizable, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot slightly shorter than tibia (96%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not recognizable. Webbing formula: 1(0.5), 2i (1), 2e(0.5), 3i (2), 3e(1), 4i (2), 4e(2), 5(0). Relative length of toes: I<II<V<III<IV. Skin on the upper surface smooth, slightly glandular dorsolaterally. Ventral side smooth. Femoral glands large and distinct in external and internal view.
Colour in preservative (after 16 years in preservative) dorsally brown, with indistinct and irregular darker markings. A dark brown band present between eyes. Forelimbs light brown with 4–5 dark brown crossbands on hand and arm. Hindlimbs light brown with distinct dark brown crossbands. Inguinal region without few scattered whitish spots. Snout tip with a light spot (width 1.3 mm, height 1.6 mm), tympanum region distinctly lighter than remaining head sides. Venter beige, throat darker than belly with brown mottling. A distinct longitudinal white median line runs from snout tip over the entire throat to the anterior thorax. Lower lip with distinct alternating light and brown spots. Toe discs dark.
Colouration in life ( Fig. 4 View FIGURE 4 ) similar to that in preservative, but more contrasting: The darker areas on the dorsal surface were more distinct; a white spot was present in the upper lip area between eye and tympanum and whitish spots were present on the lateral body. The tympanum region was distinctly darker than the remaining head side.
Description of a female paratype ( allotype). ZSM 825/2003, adult female in good state of preservation. For measurements, see Table 1. Body rather stout. Head wider than long. Head as wide as body. Snout rather pointed. Nostrils directed laterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis weak, slightly concave. Loreal region weakly concave. Tympanum distinct, small, rounded, diameter 71% of eye diameter. Supratympanic fold distinct, beginning straight, with rather distinct bend midway towards insertion of forelimb. Tongue ovoid, distinctly posteriorly bifid. Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded. Subarticular tubercles single. Outer metacarpal tubercle recognizable, inner metacarpal tubercle present. Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Foot slightly shorter than tibia (94%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle not present. Webbing formula: 1(0.5), 2i (1), 2e(0.5), 3i (2), 3e(1), 4i (2), 4e(2), 5(0.5). Relative length of toes: I<II<III<V<IV. Skin on the upper surface smooth. Ventral side smooth. Femoral glands small but present. In preservative, dorsum brown, with indistinct irregular darker markings. A dark brown and more or less triangular band between eyes present. Forelimbs lighter brown with darker markings. Hindlimbs lighter brown with distinct darker crossbands. Inguinal region without few scattered whitish spots. Snout tip without a light spot. Venter beige, throat darker than belly with brown mottling. A longitudinal indistinct white median line on thorax and throat is present. Lower lip without distinct alternating light and brown spots. Toe discs dark. Toes light and dark striped. Typical colour pattern on the side of the head is indistinctly present.
Variation. Measurements of nine available paratypes (two males and seven females) are shown in Table 1. Overall males were smaller than females (SVL 21.4–23.5 vs. 24.7–28.9 mm) and had larger tympani (TD 91–94% vs. 71–82% of ED). Dorsal colouration is almost uniformly light brown to dark brown in most paratypes with almost no difference between dorsal and lateral colouration, but in most of them a dark band between the eyes is recognizable or even distinct and a few individuals (ZSM 652/2001 and ZSM 823/2003) show a relatively well recognizable dark blotch on the central back. In contrast to the holotype, most paratypes have no white spot on the snout tip, but a small white spot is also present in ZSM 821/2003. Dark crossbands on the dorsal surfaces of the hands, arms, feet and limbs are well recognizable or even distinct in all paratypes and most individuals have a few small whitish dots along the posterior flanks (not recognizable in ZSM 821/2003). The tympanum is either as dark as the surrounding head sides (e.g. ZSM 826/2003) or distinctly lighter (e.g. ZSM 824/2003). The belly ranges from uniformly light grey (ZSM 825/2003) to dark grey (ZSM 821/2003). The throat is brown and darker than the belly, usually with small whitish dots and a thin median line, which however is always less distinct than in the holotype.
Available earlier names. As discussed below, there are numerous junior synonyms of Brygoomantis species, listed by Blommers-Schlösser & Blanc (1991). In most cases the identity of these is in need of confirmation, and many will probably refer to candidate species as defined by Vieites et al. (2009) and Perl et al. (2014). The majority of the name-bearing types of these species have larger body sizes than Mantidactylus schulzi and therefore can be excluded as available earlier names of this species. However, we advocate for the distinctiveness of M. schulzi based on its restricted geographic range in Manongarivo and Tsaratanana, a region from which no collections have become available before the 1970s, thus postdating the description of the relevant junior synonyms in Brygoomantis . We therefore exclude any of these earlier names as possibly referring to M. schulzi .
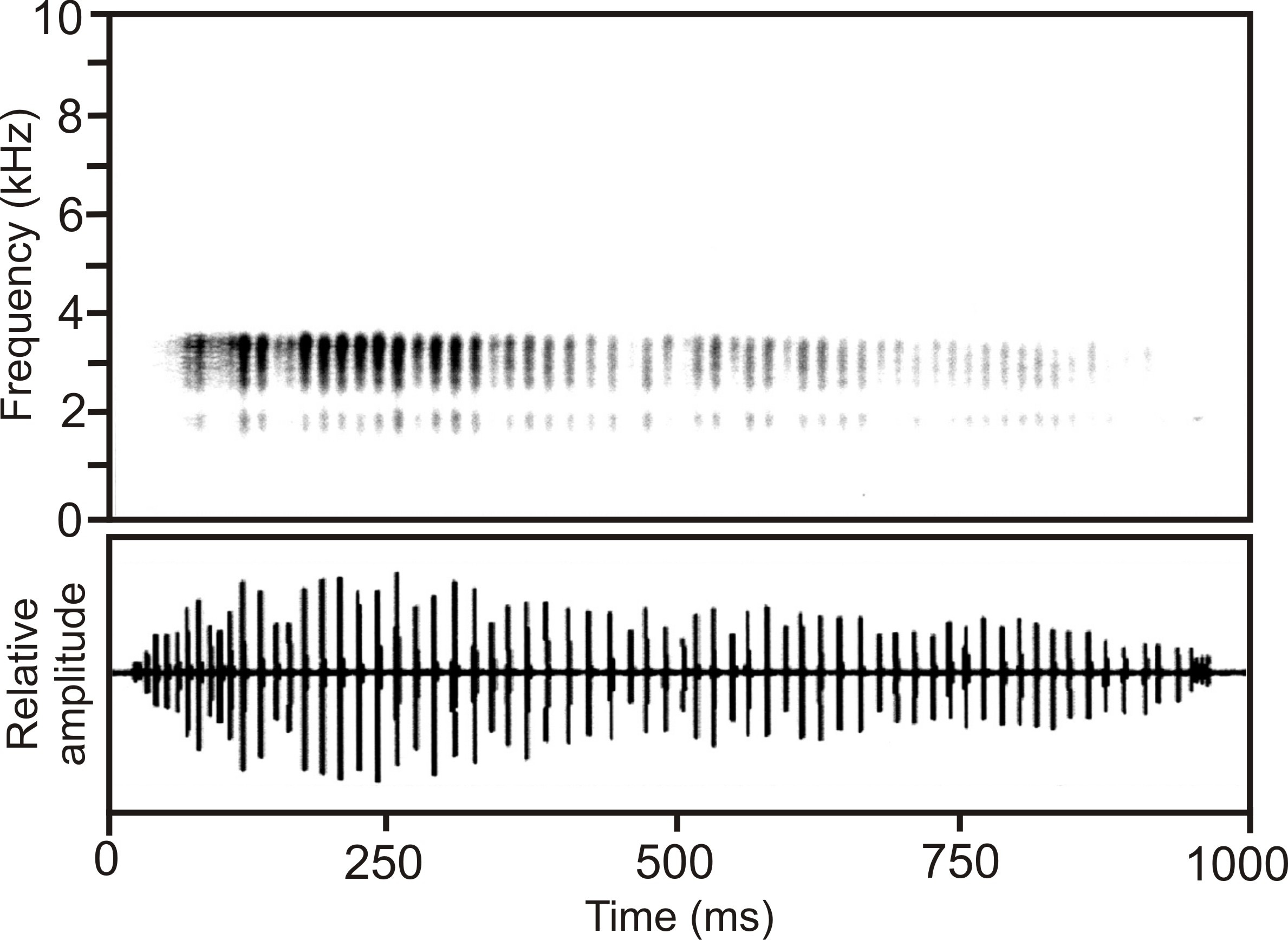
Calls. Advertisement calls were recorded by MV on 10–11 February 2001 at Andampy Campsite, Manarikoba Forest, Tsaratanana Strict Nature Reserve, 25–26°C air temperature. Calls ( Fig. 5 View FIGURE 5 ) consist of pulsed notes which sometimes are arranged in short series of 2–3 notes, with very different note duration, and different number of pulses per note; both of these variables increase from the first to the last note. Averaged over all recorded notes, these consist of 6–73 regular pulses (mean 27 ± SD 21, n=30) which last for 1–3 ms (mean 2 ms ± SD 1 ms, n=40). Note duration is 11–996 ms (mean 301 ms ± SD 306 ms, n=30). Interval duration between the notes is very irregular, spanning from 35–2716 ms (mean 673 ms ± SD 717 ms, n=28). The interval duration between the pulses is 1–18 ms (mean 11 ms ± SD 4 ms, n=40) and increases with increasing note duration. Pulse repetition rate is 62– 84/s in longer notes. Dominant frequency is difficult to determine and is distributed over a rather large range between 1900–3400 Hz; prevalent bandwidth is between 1500 and 3900 Hz.
Distribution and habitats. The species is known from low- to mid-elevations in the Manongarivo and Tsaratanana Massifs, specifically from Camp 0 at Manongarivo, and Andampy (Camp 0) in the Manarikoba Forest at Tsaratanana, at an elevational range of approximately 690–730 m above sea level. All individuals were observed along streams in rainforests.
Etymology. The new species is a patronym dedicated to Stefan Schulz, in recognition of his outstanding contributions to our understanding of pheromonal communication in anurans, and especially in species of Brygoomantis .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |