
Parartemia mouritzi
|
publication ID |
https://doi.org/ 10.5281/zenodo.199709 |
|
DOI |
https://doi.org/10.5281/zenodo.6200946 |
|
persistent identifier |
https://treatment.plazi.org/id/5D556922-6456-654F-FF0B-A67DFC50FCBF |
|
treatment provided by |
Plazi |
|
scientific name |
Parartemia mouritzi |
| status |
|
Parartemia mouritzi View in CoL
(Figs. 4,7,10)
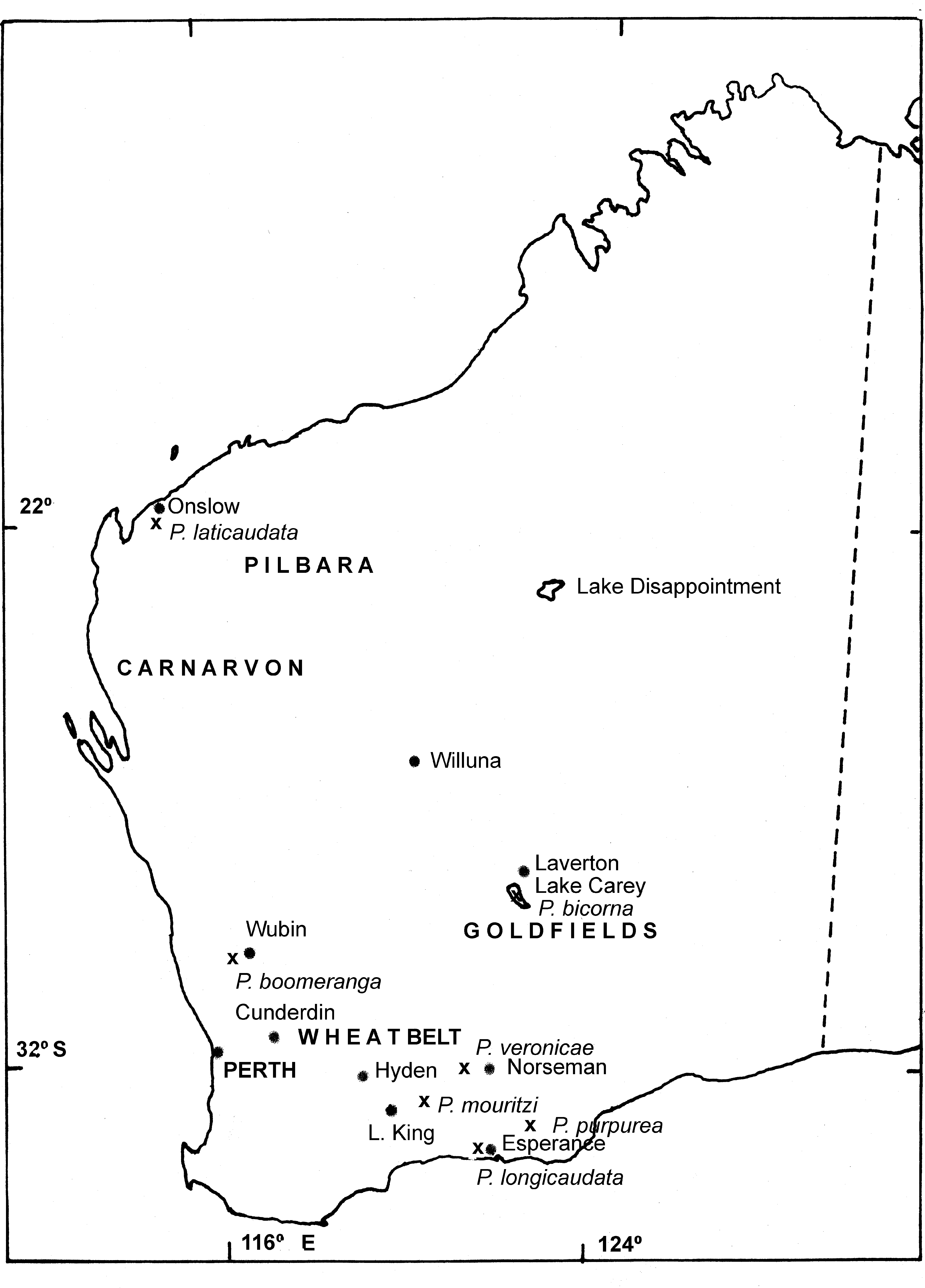
Parartemia sp b, Timms and Savage, 2004, p 20,21,31.
Etymology. This species, found in the Hyden area, is named to honour the late Russel Mouritz, a district pioneer’s son, who loved and understood the bush. He was a community leader, tourism founder and guided thousands with enthusiasm at Wave Rock. Moreover Russel loved learning about the environment, whether through his own forsightful actions on degraded land or by supporting geomorphological and biological research on Wave Rock and surrounding areas.
Holotype. Male, about 67 km east of Lake King township, unnamed claypan in Frank Hahn National Park, (32o 57’ 43.5”S, 120o 21’ 54.1”E) 7 October, 2008, B.V. Timms, WAM 45229.
Allotype. Female (ovigerous), same collecting data as holotype, WAM 45230.
Paratypes. Two males, two females, same collecting data as holotype, WAM 45231; Two males, two females, same collecting data as holotype, AM P82975.
Other material. 16 males, 6 females, 29.5 km E of Hyden, unnamed lake on Lake Carmody Rd, (32o 27’ 44S, 119o 10’ 30”E), 12 August 2008, B V Timms, WAM 45232; 20 males, 10 females, same site, 12 October, 2009, J. O’Connor & B.V.Timms, WAM 45233; 20 males, 2 females, 29.1 km E of Hyden, unnamed lake just S of Carmody Rd, (32o 27’ 54”S, 119o 10’ 15”E), 12 August 2008, B.V. Timms, WAM 49234; 3 males, 3 females, 50 km NW of Norseman, unnamed pool, (31o 50’ 36”, 121o 27’ 50”) 26 June 2010, B. Datson.
Description. Male. Length 10.8 mm (head + thorax 4.8 mm, abdomen 6.0 mm).
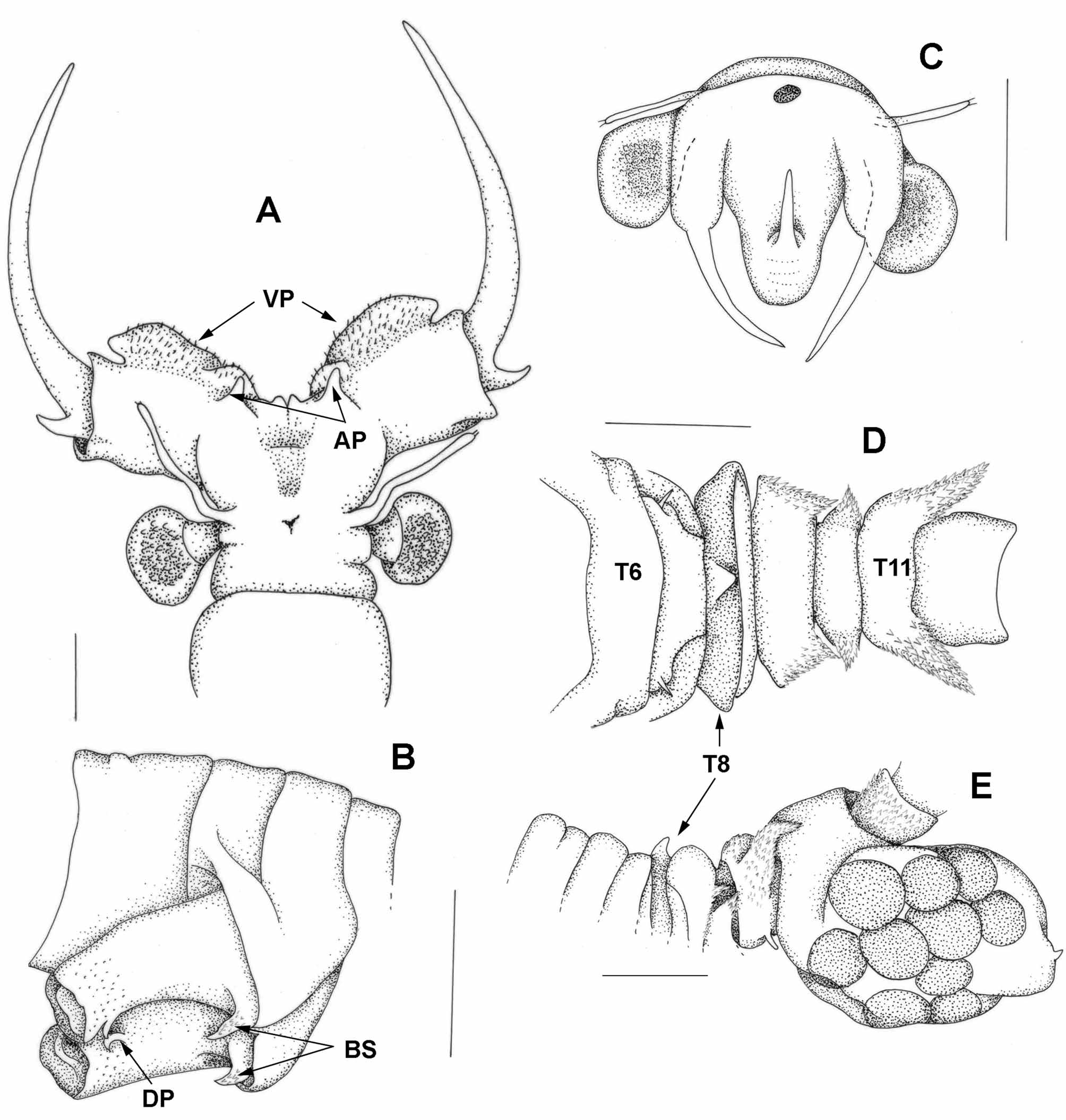
First antenna ( Fig 10 View FIGURE 10 A) filiform, slightly longer than eye plus peduncle.
Second antenna. Basal antennomere ( Fig 10 View FIGURE 10 A) of second antenna fused proximally at an angle of about 30o to body axis (but spread apart in Fig 10 View FIGURE 10 A). Ventral margin with paired, linear ventral processes ( Fig 10 View FIGURE 10 A), clothed apically with numerous small denticles mainly on medial three quarters of its length. Each process with a subsidiary process medially, and another just anteriorly and about one third the size; so a major and minor ventral process on each side. Lateral edge of major process rounded and protruding and ventral edge convex. Overall length of the two processes together about 4 times longer than high. Medial space between processes about half length of processes and in a different, more anterior, plane to the ventral processes and with a small central notch flanked by a minor protrusion on each side. Small digitiform anterior processes ( Fig 10 View FIGURE 10 A) protruding a little over the minor ventral processes. Medial surface of fused basal antennomeres with a well-defined squarish depression open ventrally. Distal antennomere of second antenna ( Fig 10 View FIGURE 10 A) about twice the length of basal antennomere and tapering to a sharp apex; a recurved spine basiolaterally, a little shorter than the diameter of the distal antennomere at its base. Polygonal pattern on surface of much of distal antennomere.
Thoracic segments unremarkable, lacking noticeable lateral lobes or projections elsewhere. Eleven pairs of thoracopods, with the first and last two pairs noticeably reduced in size.
Fifth thoracopod similar to that of P. longicaudata : endites 1+2 with about 58 posterior setae, endite 3 with 15, and endites 4, 5 and 6 with 3, 2 and 1 respectively. Basalmost anterior setae of endite 1+2 longer than posterior setae, and similarly for endite 4; other anterior setae of normal comparative lengths for Parartemia . Posterior setae with sparse setules while some anterior setae (on endite 1, 4, 5 and 6) double pectinate and others (on endites 2, 3) naked. Endopodite squarish with only 3 posterior setae (with a single pectin) on medial surface and about 15 on apical margin. Exopodite elongate oval with about 21 marginal posterior setae. Endopodite and exopodite setae based with a coronet of up to 15 small spines. Epipodite oval and praepipodite elongate oval; both unadorned. Eleventh thoracopod lacks an epipodite.
Gonopods ( Fig 10 View FIGURE 10 B) paired, basal parts fused together and about twice diameter of tubular free apical part. Base of free apical part bears a sharp spine about twice as long as thick at its base, while apical part with a short digitiform process on mediolaterially surface subapically.
Abdominal segments increasing in length and narrowing posteriorly from 1 to 6, particularly 5 and 6, so that last segment about twice the length of the first. Cercopods fringed with long setae on lateral and medial edges; length subequal to 6th abdominal segment.
Female. Length 6.2 mm (head + thorax 3.9 mm, abdomen 2.3 mm).
Head ( Fig 10 View FIGURE 10 C) with first antenna filiform, subequal in length to eye plus peduncle.
Second antenna with thick basal portion and arising symmetrically from it, a narrow apical portion a little longer than it. Prominent naupliar eye midway between compound eyes. Labrum terminating in a narrow recurved spine.
Posterior thoracic segments ( Fig 10 View FIGURE 10 D,E) with complex lateral lobes and/or dorsal protuberances. Thoracomere 6 with a small dorsal tumidity and a normal lateral lobe; thoracomere 7 also with a small dorsal tumidity but with a weak triangular lateral lobe; thoracomere 8 with a small asymmetrical lobe, swollen posteriorly terminating in a posteriorly directed apex and with a small sharper dorsal protuberance than thoracomeres 6 and 7; thoracomere 9 with a large rounded dorsal swelling and the biggest lateral lobe, again asymmetrically swollen posteriorly and terminating in a posteriorly directed apex; thoracomere 10 with a rounded lateral lobe, and sunken dorsum without any swelling; and thoracomere 11 with an increasingly elevated spinose dorsal surface and posteriorly a large lateral curved spine. This spine plus lateral lobes of segments 9 and 10 and dorsum of thoracomeres 10 and 11 sclerotized. Also, the spine together with the lateral edges of lateral lobes of thoracomeres 8–11 with denticles, particularly apically. Brood pouch not bilobed, deep (i.e. maximum dimensions at right angles to body axis) and with pigmentation over the eggs dorsally.
Ten pairs of thoracopods, first and last considerably reduced. First thoracopod (Fig 7H) of different proportions to standard Parartemia thoracopod, the endopodite almost rounded, the exopod distinctly smaller than in T2–7 and oval shaped, the epipodite oval shaped and the praeepipodite roughly rectangular. Posterior setae variable in number, with fewer on endopodite and exopodite than usual (i.e. 11 cf 15 and 14 cf 21), about the same on endites 1+2 and 3) and none on endites 4 to 6. Anterior setae of endite 1 particularly stout (basal width equal to basal diameter of two posterior setae and with strong spines, while basal most anterior setae of endite 4 elongated to exceed posterior setae, bent and double pectinate.
Tenth thoracopod different again (Figs 7I,); both with considerably reduced endites 1+2 and 3 bearing altogether few (<15) posterior setae, but endites 4 to 6 with normal anterior and posterior setae. Endopodite small (just bigger than epipodite of 9th), roundish and bearing 10–12 posterior setae in each, and epipodite lanceolate, a little longer than endopodite and with 8–10 posterior setae. Thoracopod 9 with normal epipodite and prepipodite, but thoracopod 10 lacking both. No 11th thoracopod.
Abdominal segments papillate, otherwise proportions as in male.
Variability. Parartemia mouritzi sp. nov. is small, males ranging from 7.2 to 10.8 mm and females from 4.9 to 6.5 mm. This makes it even smaller than P. minuta (males 7.4 – 13.6 mm, females 4.9 – 7.8 mm, both sets of measurements without cercopods which would add about 1 mm) ( Geddes, 1973).
In males fusion of the basal antennomeres varies from about 15 – 40o, perhaps an artefact associated with torsion during preservation. The angle never approaches 60 – 90o seen in most species of Parartemia . In some individuals the first antennae about are subequal to the eye plus peduncle rather than being longer.
In females thoracic segments 6–11 vary slightly in appearance due to incompletely developed swellings and lateral lobes. This is usually seen in females with undeveloped brood pouches, so may be associated with age. However the lateral lobe of segment 11 is variously shaped in mature females: generally it takes the form of a curved posteriorly pointed spine, but it may be more triangular and pointing laterally.
Differential diagnosis. Parartemia mouritzi sp. nov. is most similar to P. m i n u t a. Males of both species have the angle of fusion of the basal antennomere <30o, an elongated, convexly surved ventral process, ventral process with a subsidiary ridge (the minor ventral process) anteriomedially, a basolateral spine on the distal antennomere, a short digitiform process, and a straight, pointed proximal spine on the gonopod. They differ in only a few morphological characters ― the minor ventral process is denticulate in P. m o u r i t z i sp. nov. but smooth in P. m i n u t a, and the medial ventral margin of the fused basal antennomeres is shaped in the form of a broad triangular notch in P. minuta , whereas in P. mouritzi sp. nov. the notch is short with each side appressed and with a small bulge lateral to the notch. There is also a subtle difference in the basal spine of the distal antennomere; in P. minuta it is generally blunt and significantly less than the basal width of the distal antennomere whereas in P. mouritzi sp. nov. it is sharp and only a little shorter than the basal width of the distal antennomere.
Females of the two species are also similar, in that they are both small, have a single ventral brood pouch, and have dorsal projections on thoracomeres 6 and/or 7. This projection is spine-like in P. minuta and a more rounded bulge in P. mouritzi sp. nov. More distinctive is the lateral compression of segments 8–11 and lack of lateral lobes in P. minuta as against their normal comparative width and presence of lateral lobes in P. mouritzi sp. nov., that of the 11th thoracomere being particularly distinctive in its denticulated curved spine-like structure projecting posteriorly, or occasionally laterally.
Females of P. cylindrifera also bear some resemblance to those of P. mouritzi sp. nov., mainly in the sharing of a single ventral brood pouch, but again the structure of segments 8–11 is very different in the two species in that P. cylindrifera lacks the lobes of P. m o u r i t z i sp. nov. Moreover thoracopod is vestigial in P. cylindrifera whereas in P. mouritzi sp. nov. it is reduced as normal for the genus.
Distribution and ecology. P. m o u r i t z i sp. nov. is known from only three small shallow salinas in the Hyden-Lake King area of the Wheatbelt ( Fig. 4 View FIGURE 4 ). Given this restricted distribution and the secondary salinisation of two of them, Timms et al. (2009) recommend it be given Priority One status under the Western Australian Department of Conservation’s priority species listing process.
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |