
Parartemia bicorna, Timms, Brian V, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199709 |
|
DOI |
https://doi.org/10.5281/zenodo.6200940 |
|
persistent identifier |
https://treatment.plazi.org/id/5D556922-644F-6549-FF0B-A09DFC50FED9 |
|
treatment provided by |
Plazi |
|
scientific name |
Parartemia bicorna |
| status |
sp. nov. |
Parartemia bicorna View in CoL sp. nov.
(Figs. 4,7,8)
Parartemia View in CoL sp x, Timms et al., 2006, p 181, Table 5; Timms and Hudson, 2009, p 58.
Etymology. The specific epithet, bicorna , celebrates the prominent twin anterior processes of the male, vaguely akin to the head spine of the fabled unicorn, but with two instead of one horn. These process are somewhat larger than in other known species of Parartemia ; some like P. contracta may have longer processes, but these are generally narrow and bent, while those in P. b i c o r n a sp. nov. are straight and robust, and hence instantly noticeable.
Holotype. Male, Lake Carey, 27.5 km south of Laverton, Wallaby Mine discharge, (28o 52’ 03”S, 122o 19’ 25”E), 11 March 2004, B. Timms, WAM 45214.
Allotype. Female (ovigerous) same collecting data as holotype, WAM 45215,
Paratypes. Two males and two females, same collecting data as holotype, WAM 45216; two males and two females, same collecting data as holotype, AM P82973.
Other material. Seven males, six females, Lake Carey, ‘Salinaland’ (ie.northeast corner), (28o 49’ 58”S, 122o 16’ 16”E), 11 March 2004, B.V. Timms, WAM 45217; ten males, ten females, Lake Carey, Causeway area, (29o 18” 53”S, 122o 34’ 05”E), 17 February 2006, B. Datson, WAM 45218; seven males, eight females, Lake Carey, inlet into ‘Salinaland’, (28o 48’ 42”S, 122o 15’ 6”E), B. Datson, WAM 45219; nine males, 2 females, Lake Carey, ‘Salinaland’ (ie.northeast corner), (28o 49’ 58”S, 122o 16’ 16”E), 6 December 2008, B. Datson, WAM 45220.
Description. Male. Length 19.8 mm (head + thorax + genital segments 8.3 mm and abdomen 11.5 mm). First antenna ( Fig 8 View FIGURE 8 A) filiform, about twice as long as eye plus peduncle.
Second antenna. Basal antennomere of second antenna ( Fig 8 View FIGURE 8 A) fused proximally at about right angles to body axis and protruding a similar width to first antenna. Ventral margin of second antenna with paired linear ventral processes ( Fig 8 View FIGURE 8 A) bearing numerous closely spaced minute denticles apically. Overall dimensions of these processes about 2.5 times longer than average width, with lateral edge about half length of medial edge. Lateral edge broadly rounded but medial edge folded anteriorly to about one fifth of ventral process length; fold widest at about two-thirds length of the ventral process. Medial edge of ventral process bulges into the medial space between the processes, maximally at about half its height. Posterior base of lateral edge of ventral process with an indistinct conical mound with a few small denticles. Medial process digitiform, length slightly less than half the height of the ventral process medial edge. Anterior surface of proximal antennomeres with a prominent conical anterior process ( Fig 8 View FIGURE 8 A) on each half, with a large free apical half about the length of the ventral process. Distal antennomeres ( Fig. 8 View FIGURE 8 A) of second antenna about twice length of basal antennomere, curved, with a mesial tumidity and tapering to a sharp apex. Labrum lacking a spine.
Thoracic segments ( Fig 8 View FIGURE 8 B) with lateral lobes progressively increasing in size T1 to T11 and then with smaller lobes on the two genital segments. Eleven pairs of thoracopods, variable in size, with first two noticeably reduced in size and last few a little smaller. Fifth thoracopod similar to that of Parartemia longicaudata . Endite 1+ 2 with 1+2 anterior setae and ca. 25 posterior setae, endite 3 with 2 and ca 16 setae respectively, endite 4 with 2 and 3, endite 5 with 2 and 2 and endite 6 with 1 and 2 setae respectively. These setae of similar relative lengths to those of P. acidiphila ( Timms and Hudson, 2009, Fig 2). Endopodite squarish with broadly rounded corners and bearing ca, 6 one-sided pectinate posterior setae medially and ca. 24 feathered posterior setae terminally and laterally. Exopodite elongate oval and bearing ca. 36 posterior setae, similar to most on the endopodite. All these posterior setae with a basal coronet of ca. 8 small spines. Epipodite oval shaped and praeepipodite elongated oval shaped, both unadorned.
Gonopods ( Fig 8 View FIGURE 8 C) paired, basal parts fused together and about twice the diameter of apical free portions. Free apical part basally with a triangular process with an asymmetrical apex and a subapical digitiform process, about a third the length of the basal process. No type specimens with the gonopod everted.
Abdominal segment s increasing in length and narrowing posteriorly from 1 to 6, particularly 5 and 6, so that last segment about twice the length of the first. Cercopods fringed with long setae on lateral and medial edges; length subequal to 6th abdominal segment.
Female. Length 14 mm (head + thorax+ genital segments 7.2 mm, abdomen 6.8 mm).
Head ( Fig 8 View FIGURE 8 D) with first antenna filiform and length subequal to eye plus peduncle. Second antenna subcylindrical but with a markedly narrower recurred apex; antenna a little longer than eye plus peduncle. Naupliar eye distinct and midway between compound eyes. Labrum with a prominent recurved spine.
Thorax ( Fig 8 View FIGURE 8 E) without dorsal or lateral tumidities, but thoracic segment 9 with a prominent lateral lamellar lobe extending posteriolaterally to segments 10 and 11. Segments anterior to ninth with small lateral lobes, biggest on segment 8. Lateral and posterior edges of the segment 9 lamellar lateral lobe sclerotized; dorsum of segment 11 also sclerotized and with a minor round protuberance laterally. Brood pouch lying laterally to the genital segments, each with anterior edge sclerotized, rounded and adpressed against the posterior edge of the lateral lamellar lobe of segment 9. Each brood pouch half pyriform and joined ventrally to a gonopore on a short tubular process. Eggs round and smooth-surfaced at low magnifications.
Ten thoracopod pairs, with first half the size of the others and the last three a little shorter. Eleventh thoracic segment without appendages. Thoracopods 2–7 similar to those of male though proportions slightly different, most noticeably the broader than long endopodite. First thoracopod (Fig 7C) of very different proportions than in standard Parartemia thoracopod. Endopodite almost round, exopodite small and oval shaped, epipodite oval shaped and praeepipodite sausage shaped. Endopodite and exopodite with fewer posterior setae than usual, namely about 14 and 20 respectively. Endites 1+2 and 3 of normal relative size and setation, but endites 4 to 6 very different; all three lack posterior setae and 4 and 5 bear elongated bent anterior setae with apical twofifths with a double pectin. Thoracopod 10 (Fig 7D) is different again, lacking the epipodite and praeepipodite and with a reduced rounded endopodite (posterior setae reduced to 2 on medial surface and ca. 12 on terminal and lateral edges), but exopodite more typical (and with ca 24 posterior setae). Endites 1+2 and 3 bear typical anterior setae, but few posterior setae (ca. 12 and 3 respectively). Endites 4 to 6 without posterior setae and anterior setae reduced to one each with those of endite 4 and 5 longer than usual.
Abdominal segments papillate, otherwise proportions as in males.
Variability. Body lengths in the study material vary up to 20.4 mm in males and 14.2 mm in females. Fusion of the basal antennomeres is sometimes not at right angles to body axis, but as low as 75o. The distinctive frontal processes are only fully developed in mature males. The fold on the medial edge of the ventral processes sometimes has a distinct point mid length and reaching almost to the ventral edge of the process. The ventral conical mound at the base of the lateral edge of the ventral processes is indistinct in many specimens.
The main variability on females concerns the area between the lateral edge of segment 9 and the forward extension of the brood pouch: in some specimens there is a hook-like structure pointing forward or at least a small outgrowth laterally. In a few specimens the dorsal integument of segment 11 is only weakly sclerotized. The second antennae do not always terminate in a thin curved point, but may be more mamilliform.
Differential diagnosis. Male P. bicorna sp. nov. is most similar to males of P. serventyi and to a lesser extent to males of P. informis . Distal antennomeres in both P. b i c o r n a sp. nov. and P. serventyi are identical, both with a characteristic mesial tumidity. Anterior, ventral and medial processes are almost identical in the two species, but there are three minor differences: (a) the anterior processes in mature P. b i c o r n a sp. nov. are more robust, being 1.25: 1 longer than wide, whereas in P. serventyi the ratio is closer to 1.5:1, (b) the fold on the medial edge of the ventral processes is more pronounced in P. bicorna sp. nov., and (c) the ventral conical mounds on the base of the lateral margin of the ventral processes, if present, are small compared with those in P. serventyi which are larger and separate from the ventral processes. There is a more obvious difference between the two species in the lateral lobes of the thorax and genital segments―those of P. b i c o r n a sp. nov. show a gentle gradient in size with lobes of the 11th thoracic segment just the greatest, whereas in P. serventyi , the lobe on the 11th thoracic segment is at least twice the size of that of the 10th segment or first genital segment.
The difference between P. b i c o r n a sp. nov. and P. informis is more pronounced. The anterior processes are much shorter in P. informis than in P. bicorna sp. nov., and the ventral processes lack a medial fold in P. informis compared to a fold being present in P. b i c o r n a sp. nov. Two minor differences are in the lateral corner of the frontal processes which is rounded in P. b i c o r n a sp. nov. but square in P. informis and in the number of spines in the basal coronet of the endopodal setae: <10 in P. b i c o r n a sp. nov. and> 20 in P. informis . Also there are more anterior setae on the endites 1 to 3, the endopod and the exopod in P. informis , but this could be because P. informis is generally larger (often> 30 mm) than P. b i c o r n a sp. nov. (usually <20mm).
Males of P. bicorna sp. nov. also resemble the males of P. contracta to some degree in that the anterior and ventral processes are broadly similar. However the anterior processes are usually thin and bent in P. contracta compared to being stout and triangular in P. bicorna sp. nov. and the ventral processes lack a medial fold in P. contracta . Another difference is the lack of a mesial tumidity in the distal antennomeres of P. contracta .
Among females there can be no confusion between P. b i c o r n a sp. nov. and P. serventyi . The later has a marked medial bulge on the dorsal surface of the 8th thoracic segment and no lateral plate on segment 9, whereas P. b i c o r n a sp. nov. has no bulges on segment 8 and has a lateral plate on segment 9. Likewise P. contracta is different in that the lateral lobe of segment 9 terminates in a point posteriolaterally and does not envelop segments 10 and 11, whereas in P. b i c o r n a sp. nov. the lateral lobe of segment 9 envelops segments 10 and 11 and terminates in a rounded posterior edge.
Female P. bicorna sp. nov. are most similar to female P. informis . Both have a lateral lamellar lobe on segment 9 enveloping segments 10 and 11 and both have laterally bulging brood chambers. There are two lateral papillae on the dorsum in the enclosure, but on thoracomere 10 in P. informis and thoracomere 11 in P. bicorna sp. nov. The lateral extension of thoracomere 9 is wider than long in P. b i c o r n a sp. nov. and narrower than wide in P. informis . Should the specimens be appropriately preserved there is a moderate budge medially on segment 9 in P. informis , but not in P. bicorna sp. nov.
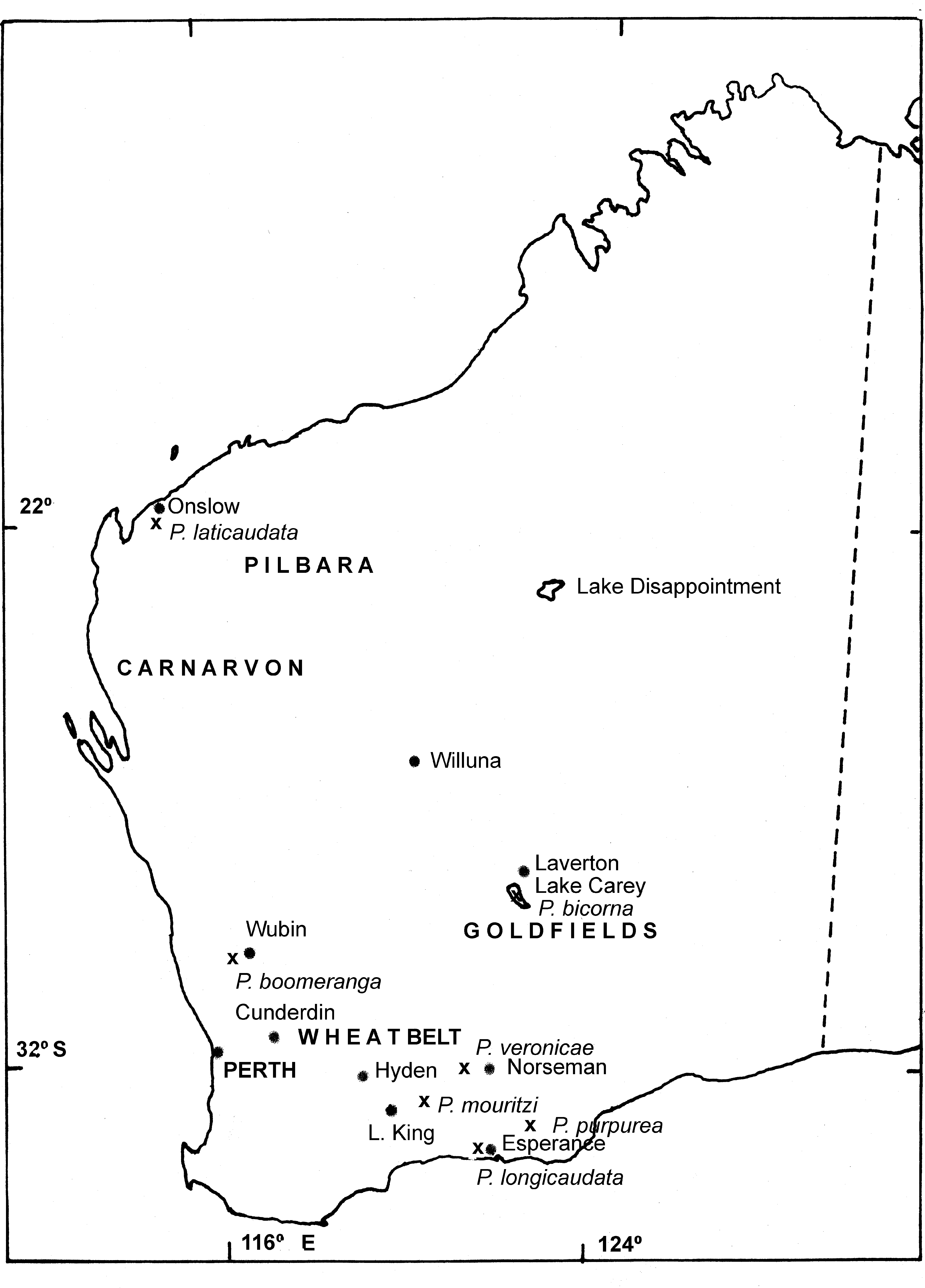
Distribution and ecology. To date, P. bicorna sp. nov. has been found only in Lake Carey, near Laverton in the northern goldfields of Western Australia ( Timms, et al., 2006, 2009) ( Fig. 4 View FIGURE 4 ). It occurs only in the main lake and sometimes shares this with Parartemia laticaudata sp. nov., an inhabitant of salinas around the lake and pools in inflowing creeks and elsewhere in northern Western Australia ( Timms et al., 2006, 2009), and also with Branchinella simplex ( Timms et al., 2006) . Field salinity range is 22–105 g /L and it occurs in any season after rain fills part or all of the lake ( Timms et al., 2006; B. Datson, pers. comm.). Given it is restricted to just Lake Carey, Timms et al. (2009) suggest it should be given Priority One status under the Western Australian Department of Conservation’s priority species listing process.

| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |