
Corethrella ( Corethrella ) kipferi Dorff, Borkent and Curler, 2022
|
publication ID |
https://doi.org/10.1080/00222933.2022.2090868 |
|
publication LSID |
lsid:zoobank.org:pub:7A1DEC81-E355-44FC-B7CD-F444663ED9CE |
|
DOI |
https://doi.org/10.5281/zenodo.7016701 |
|
persistent identifier |
https://treatment.plazi.org/id/5C6E878A-3B29-FFB8-8566-19ABF55FF927 |
|
treatment provided by |
Plazi |
|
scientific name |
Corethrella ( Corethrella ) kipferi Dorff, Borkent and Curler |
| status |
sp. nov. |
Corethrella ( Corethrella) kipferi Dorff, Borkent and Curler View in CoL , sp. nov.
( Figures 4–9 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 )
Type material
Holotype, female adult on microscope slide, labelled ‘ HOLOTYPE Corethrella kipferi Dorff, Borkent & Curler’, ‘ 16 km SE of Ozark, Missouri, 36° 52.697 ʹ N 93° 9.312 ʹ W, 280 m, 6-VI-2018, N. Dorff, frog call trap’ ( USNM) GoogleMaps . Paratypes: 1 on microscope slide from type locality, GenBank® accession number OK624783 View Materials ( USNM); 24, pinned, from type locality (6, CNCI; 8, SEMC; 10 USNM); 24, on microscope slides, from type locality (8, CNCI; 8, SEMC; 8 USNM); 5 fourth-instar larvae on microscope slides from type locality (2 CNCI; 3 USNM); 5 fourthinstar, 5 third-instar, 6 second-instar, 6 first-instar larvae, on microscope slides, locality labelled as for holotype but ‘36° 53.152 ʹ N 93° 9.271 ʹ W’’, from variable dates (4 fourthinstar, 4 third-instar, 4 second-instar, 2 first-instar, all on one slide, CNCI; 1 4 th, 1 3 rd, 2 2 nd, 4 1 st, USNM).
Diagnosis
Adult male: Unknown.
Adult female: Only extant species of Corethrella in the New World with a circular head (in anterior view, Figure 5 View Figure 5 ), with flagellomere 1 moderately elongate ( Figure 5 View Figure 5 ), sensilla coeloconica present only on flagellomeres 1, (6), 10–13 and with only a single sensillum coeloconicum on each of 10–13, wing with at least a few slightly expanded scales ( Figure 6 View Figure 6 ), with uniformly pigmented wing ( Figure 6 View Figure 6 ), scutum, katepisternum and legs ( Figure 4 View Figure 4 ). Also, the only extant species of Corethrella in the Nearctic with the following combination of characters: wing uniformly pigmented, palpal segment 3 swollen apically, and thorax nearly uniformly light brown.
Pupa: unknown.
Fourth-instar larva: only extant species of Corethrella in the Nearctic with an entirely pale body, prothoracic sensilla 7, 10 plumose, 8 bifurcate and mesothoracic sensilla 8 plumose, with four branches.
Description
Adult male: unknown.
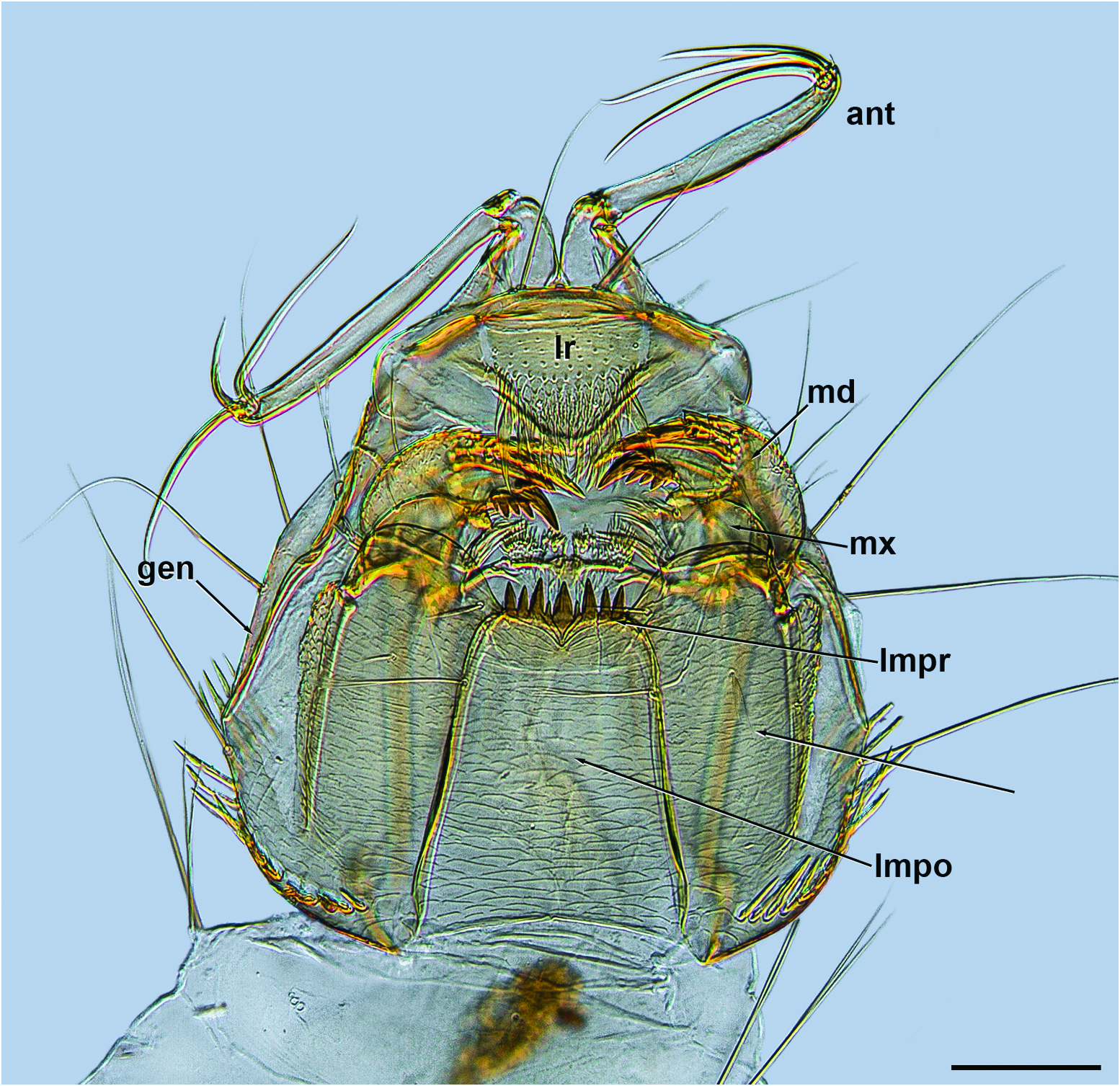
Adult female (n = 5). Head: Outline in anterior view somewhat circular (as in Figure 5 View Figure 5 ), head capsule width (W) 0.38–0.40; height (H) 0.30–0.32 W/H = 1.26–1.25. Coronal suture extending to the ventral margin of the innermost ommatidia (as in Figure 5 View Figure 5 ). One large seta on frons between ventromedial areas of ommatidia (as in Figure 5 View Figure 5 ). Antenna medium brown ( Figure 4 View Figure 4 ); pedicel ( Figure 5 View Figure 5 ) without distinctive, more elongate, stout, dorsal or dorsolateral setae; flagellomeres as in Figure 4 View Figure 4 , sensilla coeloconica distribution: 1, (6), 10– 13, with single sensilla coeloconica on each of flagellomeres 10–13; flagellomere 13 with well-developed apical bifurcation. Mandible ( Figure 5 View Figure 5 ) with small, pointed teeth. Clypeus ( Figure 5 View Figure 5 ) nearly square in frontal view, with lateral margins slightly convex, ventral margin emarginate at median, with 7–8 setae. Palpus ( Figure 5 View Figure 5 ) brown; segment 3 swollen apically, length/width (L/W) = 3–3.5.
Thorax ( Figure 4 View Figure 4 ): Nearly uniformly medium brown, small dark area on lateral margin of scutum near base of wing, pale sclerites around base of wing. Prescutal suture elongate, with area of pale cuticle at base. Posterior portion of dorsocentral row with two elongate setae situated somewhat lateral to one another. Anterior anepisternum divided diagonally by sinuous suture, dorsal portion about equal to ventral portion. Five posterior anepisternal setae. One anepimeral seta. Ventral portion of posterior anepisternum not differentiated from dorsal portion, anterior margin more darkly pigmented than the pale posterior margin.
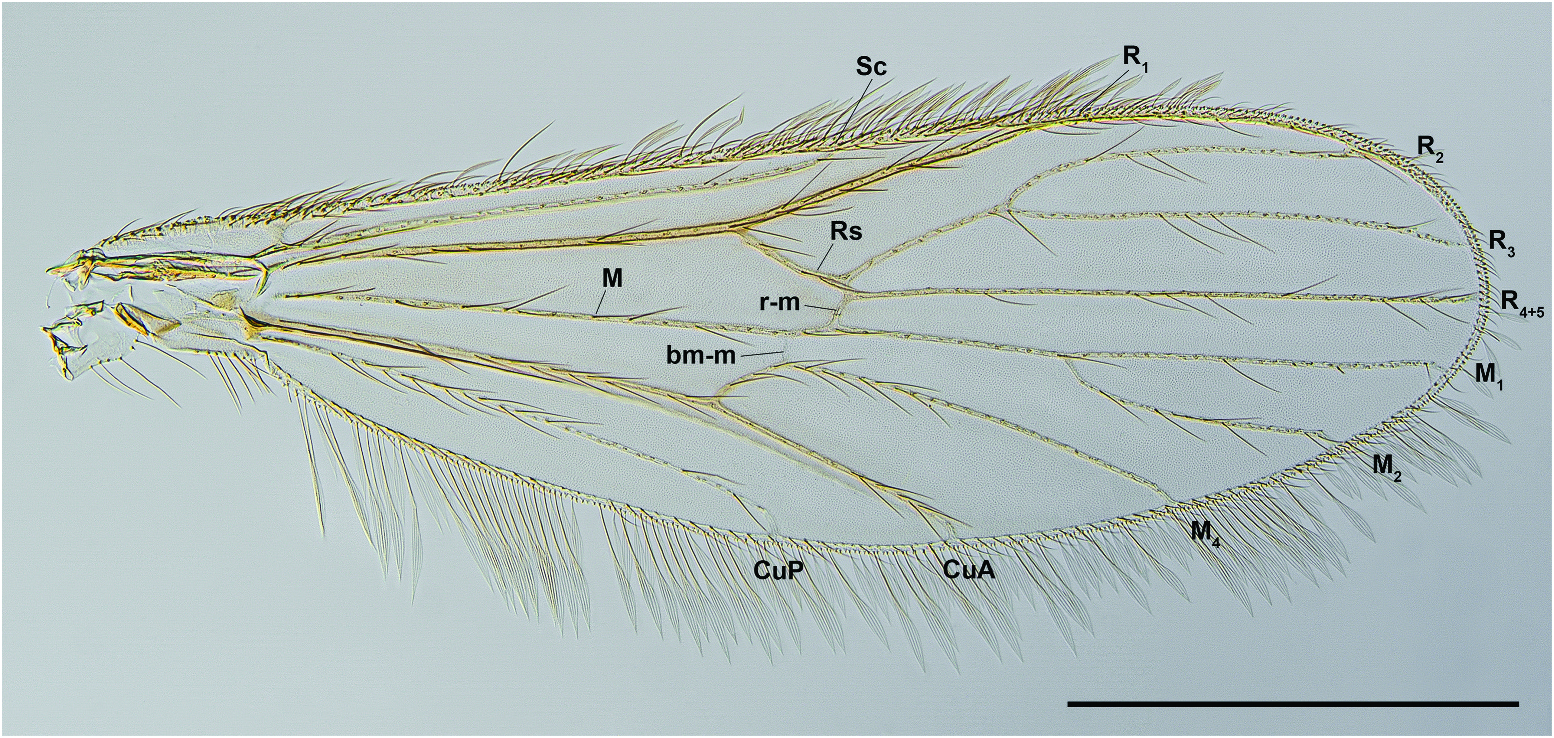
Wing ( Figure 6 View Figure 6 ): Length = 1.23–1.36 mm. Ratio of R 1 /wing length = 0.63–0.71. Apex of R 2 basal to apex of M 1. Plain, without pattern of pigmented veins and/or scales; veins (other than Costa and wing margin) with at least some slightly expanded scales. Halter paler than scutellum.
Legs (as in Figure 4 View Figure 4 ): Uniformly pigmented. With only slender setae, lacking scales (except for some in patch of whip-like setae on posterior portion of hind tibia). Mid leg with thick, subapical setae on each of at least tarsomeres 1–3. Claws of each leg equal to those of others; equal on each leg, simple (without inner teeth). Empodia slender.
Abdomen: Uniformly medium brown.
Pupa: Unknown.
Fourth-instar larva ( Figures 7–9 View Figure 7 View Figure 8 View Figure 9 ): Colouration uniformly pale ( Figure 7a–c View Figure 7 ). Head capsule L 0.52–0.60, W 0.50 –0.58, L/ W 0.92 –1.20 (n = 8); sclerites yellow to light tan, with membranous parts white; cuticle of thorax and abdomen white to transparent, with sclerites of segments 7–10 light tan. Head ( Figure 8 View Figure 8 ): Posterior margin of head capsule broadly arched anteriorly ( Figure 7c View Figure 7 ), semi-circular in dorsal view ( Figure 7a View Figure 7 ); frontoclypeal apotome approximately half the length of gena, with posterior margin clearly anterior to tentorial grooves and larval eye spots in dorsal view; labium with basal 1/3 of lateral margins slightly convex, apical 2/3 straight, converging gradually, without abrupt narrowing near apex, with apical margin (sans postmentum) appearing bilobed, cleft at median due to V-shaped furrow; postmentum with 11 dentations ( Figure 8 View Figure 8 ); median tooth largest, flanked on each side by one tooth less than half its size, three larger teeth and one minute outer tooth; outermost teeth lightly pigmented, poorly visible in cleared specimens; all teeth with bases arranged in a row so that a straight horizontal line may be placed across them in ventral view ( Figure 9 View Figure 9 ).
Thorax ( Figures 7 View Figure 7 and 9 View Figure 9 ): Prothoracic sensilla 7 and 10 plumose, 8 bifurcate; mesothoracic sensilla 8 plumose ( Figure 9 View Figure 9 ), with four branches, 13 plumose, with five branches; metathorax with lateral lobes extending beyond those of abdominal segments 1–2 ( Figure 9 View Figure 9 ), setae 6, 7, 9, 12 plumose, 10 and 13 simple.
Abdomen ( Figures 7 View Figure 7 and 9 View Figure 9 ): Dorsum of segments 1–6 lacking sclerotised plates ( Figure 7a View Figure 7 ). Lateral lobe of segments 1–2 elongate, with those of segment 2 slightly longer than those of segment 1, approximately 0.5× length of seta 6 of each segment; lateral lobes trifurcate at apex ( Figure 9 View Figure 9 ), with branches arranged in vertical row, each with elongate, simple seta inserted apically; dorsal and ventral branches of lateral lobe with minute, spiniform cuticular projections at base and elongate, prominent spiniform projections at apex, flanking base of setae alveoli; middle branch of lateral lobe with apex rounded, lacking spines. Segments 3–4 with lateral setae 6–7 plumose, each inserted on conical protuberance, setae and conical protuberances of segment 4 slightly longer than those of segment 3, both segments with lateral setae clearly shorter than corresponding simple lateral setae on segments 1 and 2. Segments 5–6 with lateral setae 5–6 plumose, decreasing in size from segment 5 to 6. Segments 6–7 with seta 13 plumose ( Figure 9 View Figure 9 ), prominent, increasing in size from segment 6 to 7. Segment 8 with lateral lobe trifurcate ( Figure 9 View Figure 9 ); ventral branch of lobe with spiniform projections at apex flanking alveolus of seta 5; seta 5 plumose, seta 4 simple. Respiratory siphon with setae 7 and 9 sharply hooked at apex, setae 7 sickle-shaped in lateral view.
Third-instar larva: Head capsule L 0.32–0.38; W 0.38 –0.40, L/ W 0.84 –0.95 (n = 5).
Second-instar larva: Head capsule L 0.24–0.30; W 0.24 –0.26, L/ W 1.08 –1.15 (n = 4).
First-instar larva: Head capsule L 0.18–0.20; W 0.18, L/ W 1.00 –1.11 (n = 4).
One female adult C. kipferi was barcoded using cytochrome oxidase subunit I (COI) and is available publicly on GenBank® (accession number: OK624783 View Materials ; www.ncbi.nlm.nih.gov/ genbank/).
Distribution
Corethrella kipferi is known only from two closely situated localities in the Ozark Mountains in Christian County, Missouri, USA, at an altitude of 280 m ( 36.92556°N 93.22528°W) GoogleMaps .
Bionomics
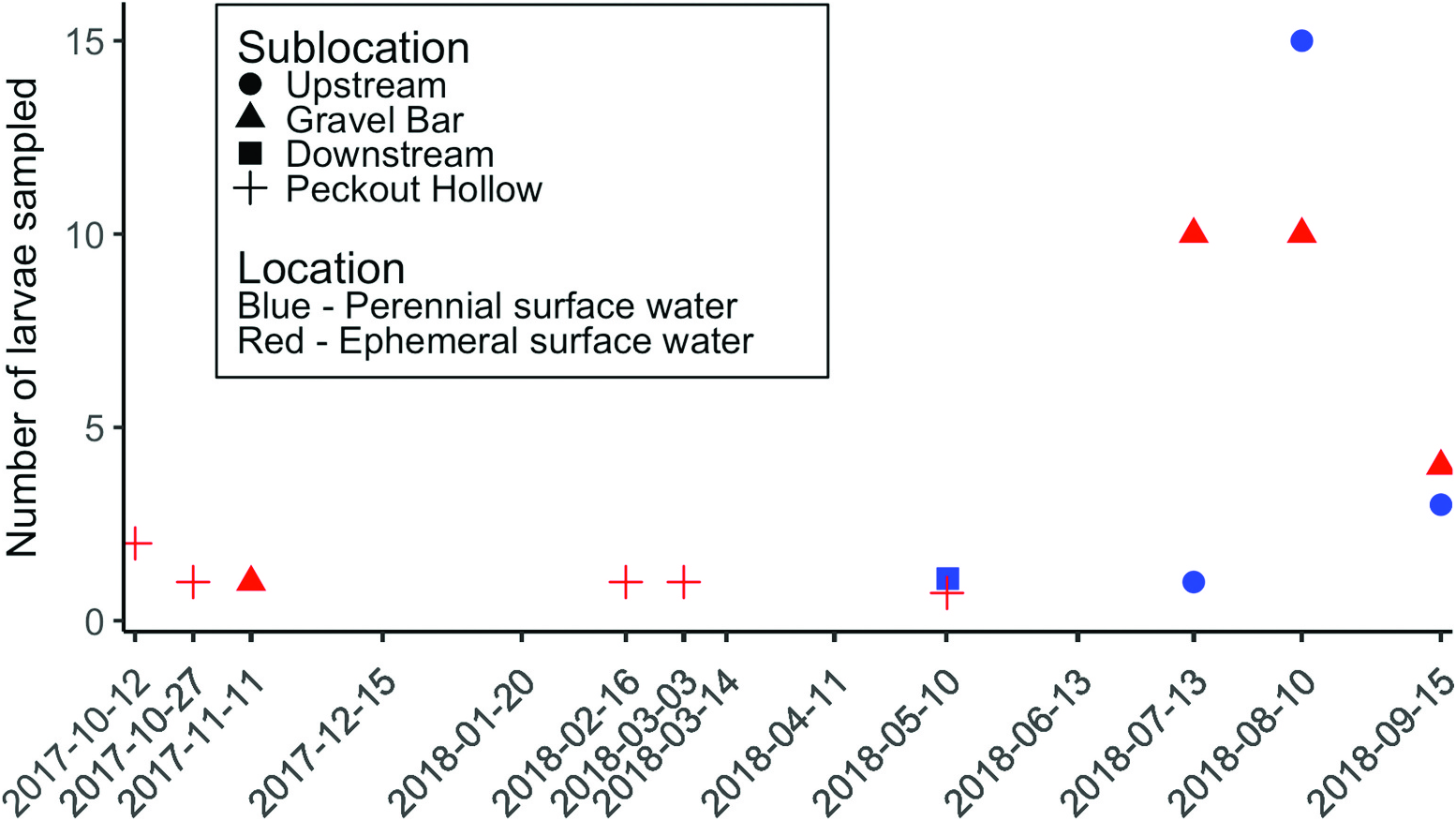
A total of 156 adult specimens were collected from a single night of trapping on 6 June 2018 using the CDC trap. In addition, seven female individuals were collected in emergence traps that were deployed on dry gravel near the stream margins. On 13 June 2018, one came from an emergence trap on the gravel bar in Bull Creek , and three individuals were taken from three different emergence traps in Peckout Hollow. On 13 July 2018, one C. kipferi adult with an abdominal blood meal was collected in Peckout Hollow, and one adult was collected in a gravel bar emergence trap in Bull Creek . On 26 July 2018, one female was collected in the gravel bar of Bull Creek . The presence of a blood-fed female clearly indicates that she was entering the trap after having fed on a frog elsewhere. No adults were detected in the emergence traps deployed upstream and downstream of the gravel bar where there was perennial flow. Additionally, we ran four emergence traps in a bedrock riffle upstream of the gravel bar reach of Bull Creek and no adult C . kipferi were collected. This may indicate the necessity of interstitial habitat for larval C. kipferi . However, due to low numbers of individuals collected in emergence traps (n = 7) there was no significant difference ( P = 0.13, df = 25) in adult capture between the gravel bar and Peckout Hollow ephemeral surface water habitats (mean = 0.538, Standard Error GoogleMaps (SE) = 0.235, 95% Confidence Interval (CI) 0.053–1.023) and the perennial Bull Creek habitats (mean = 0.00, SE = 0.235, 95% CI −0.485–0.485). Considering that adult males generally emerge, on average, earlier than adult females in most Culicomorpha , it seems likely that the presently unknown males of C . kipferi emerge earlier than the beginning of June and were missed in this study due to incomplete sampling.
Sampling of larvae suggests further possibilities. Table 1 View Table 1 gives the dates of collection of the various instars of larvae. In other Culicomorpha, first instars are of short duration and soon morph to second instars; these too moult rapidly to third instars which are of longer duration. In the majority of Culicomorpha in temperate regions, it is the last instar that overwinters ( Culicidae are an exception, with most overwintering as eggs). If males and females are mating earlier in the season, our tentative data suggests that females lay eggs in April or May (as evidenced by the single second instar recorded here) and are longer lived and still laying eggs late in the season in August (with more numerous first and second instars in August and September). If true, the females are likely multiparous and perhaps take blood meals between gonotrophic cycles, making them more likely candidates as vectors of Trypanosoma to anurans. Obviously, further research is required to understand the life cycle of this species more fully.
All C. kipferi larvae were collected with hyporheic wells. The streambed consisted of mostly gravel and cobble (median grain particle size = 32 mm; 90% CI 16–64) in both habitats. The flow in Bull Creek is perennial while the flow in Peckout Hollow is intermittent, but both channels flood following high-intensity rainfall events. One effect of the floods is the maintenance of a thick layer of well-mixed gravel (up to 8 m deep in some spaces) that increases interstitial space and delivery of dissolved oxygen and nutrients to the hyporheic zone. No significant difference ( P = 0.65, df = 24) in larval presence was detected between the gravel bar and Peckout Hollow ephemeral surface water habitats (mean = 2.21, SE = 1.02, 95% CI 0.121–4.31) and the perennial Bull Creek habitats (mean = 1.54, SE = 1.05, 95% CI −0.634–3.71).
The serrate mandibles of the female adults and their attraction to Dryophytes versicolor calls suggest that they feed on anuran blood in nature. The collection of one female with a blood meal confirms that they feed.There are 11 frog and toad species including D. versicolor known to occur near the type locality, suggesting further possible hosts ( Johnson 2000).
We attempted to rear fourth-instar larvae collected in October 2018 but were unable to get them to feed on the Caenorhabditis elegans Maupas that were offered. It is possible, given that the specimens were collected prior to spring warm-up, that they were in diapause, when feeding was minimised.
One fourth-instar larva collected from Peckout Hollow on 10 May 2018 had four head capsules in its foregut, with at least one of these belonging to a chironomid (probably Orthocladinae ), indicating in part the diet of this hyporheic larva.
Corethrella kipferi likely uses the hyporheic habitat throughout development as all instar larvae were collected only from the hyporheic zone. The larvae were also detected throughout the year in the hyporheic zone, although collected specimens were all from ephemeral surface water habitats from October through March ( Figure 10 View Figure 10 ). It is not entirely clear what types of advantages living in the hyporheic zone offers, but hypotheses include escape from predation or competition or disturbance avoidance (hyporheic refuge hypothesis; Dole-Olivier 2011).
Remarks
Our assessment of the larval chaetotaxy in Corethrella kipferi focused primarily on thoracic and ventral abdominal setae. Descriptions of shape and assignment of numbers to setae are based largely on comparison with fig. 26C in Belkin and McDonald (1955). A comprehensive survey and reassessment of the chaetotaxy in larval Corethrella overall is badly needed but is not within the scope of the present paper.
Variation in measurements of larval head capsules can occur due to several factors; however, taking consistent measurements of this structure in specimens of C. kipferi was particularly difficult due to the variation in how the mouthparts were fixed in preserved specimens. The head capsule width measured less in specimens with the buccal cavity fixed wide open than in those with the mouthparts completely retracted. This measurement can also be significantly influenced by clearing and mounting the specimen,as the lateral portions of the genae become very pliable in cleared specimens, resulting in distortion of the head capsule when placed in Canada balsam. For these reasons, it is important to measure as many ethanol-preserved specimens as possible to account for variation and it is inadvisable to rely only on measurements of slide-mounted specimens. Regardless, our measurements of head capsule length gave discrete differences between the four larval instars.
Etymology
The specific epithet, kipferi , is proposed in honour of Bob Kipfer of Springfield, Missouri, one of the landowners who allowed the first author access to their property to study invertebrates living beneath the streambed. Bob and Barb Kipfer are enthusiastic volunteers with the Master Naturalists Program of Missouri. The Kipfers also volunteer at the Wolf School, which is operated in partnership with the Springfield Public School System, Wonders of Wildlife and Bass Pro Shops and exposes youths to a variety of experientialbased programmes aimed at promoting conservation. Without the Kipfers’ permission and flexibility regarding some strange floating and even stranger frog-calling traps, we would not have been able to describe this species and its natural history with as much detail as we have. Bob and Barb Kipfer have since deeded their property to Missouri State University to be managed as a remote undergraduate and graduate research site, which will facilitate fresh insights and new discoveries in the future.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |