
Chiroleptes platycephala, (Gunther, 1873), Gunther, 1873
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4126.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:D5D8286A-E9B2-4FD8-B39A-304678C4ABD8 |
|
DOI |
https://doi.org/10.5281/zenodo.5670936 |
|
persistent identifier |
https://treatment.plazi.org/id/5230723F-0A64-FFA1-FF22-FA4F937ADEE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Chiroleptes platycephala |
| status |
|
Cyclorana platycephala ( Günther, 1873)
Eastern Water-holding Frog Figs. 5–9 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9
Chiroleptes platycephalus Günther, 1873
Holotype: BMNH 1947.2.18.42, adult female collected at Fort Bourke, NSW, Australia.
Synonymy (by Tyler 1990)
Cyclorana slevini Loveridge, 1950
Material examined. Details of the 143 specimens used to document variation are presented in Appendix 1.
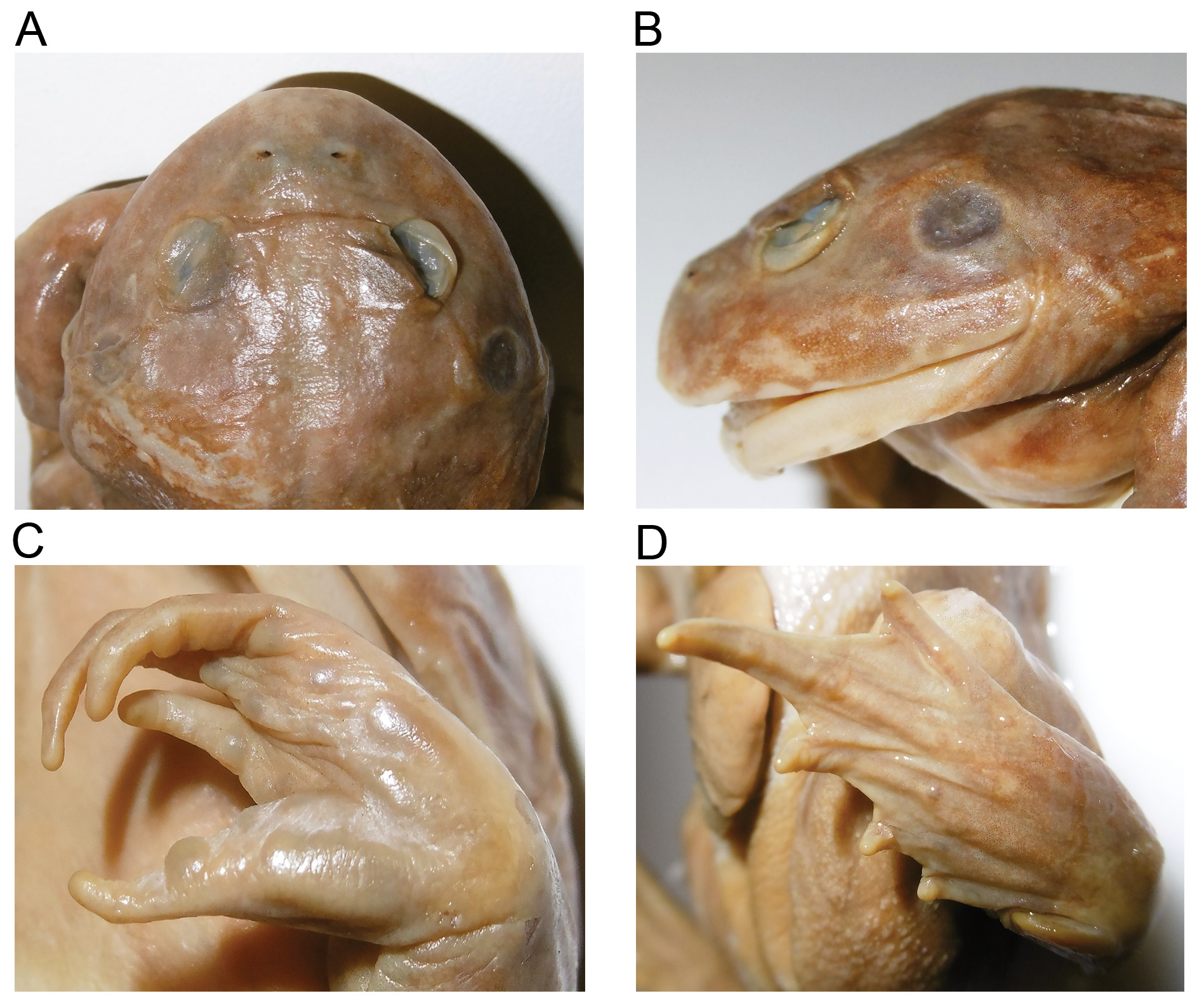
Holotype meristics. Günther (1873) presented a detailed description of the holotype but few measurements. We include images of the holotype ( Fig. 6 View FIGURE 6 ) and comprehensive set of measurements (mm): SVL = 52.9, FA = 12.8, HW = 25.5, HL 22.4, IO = 9.3, IN = 3.0, ED = 3.5, EN = 4.4, SN = 5.6.
Redefinition of Cyclorana platycephala
Diagnosis. A medium to large robust burrowing species (SVL: males 43–63 mm, females 47–72 mm). Clearly assigned to the Cyclorana by its phylogenetic placement in molecular genetic analyses ( Figs. 2 View FIGURE 2 ) and a combination of its morphological characters, burrowing behaviour, tadpoles and call structure. Distinguished from all other congeners, except C. occidentalis , by the more dorsally-tilted eyes, fully webbed toes (no more than half-webbed in all other Cyclorana), and the oral disc of tadpoles ( Anstis 2013). For distinction from C. occidentalis see Diagnosis for C. occidentalis below.
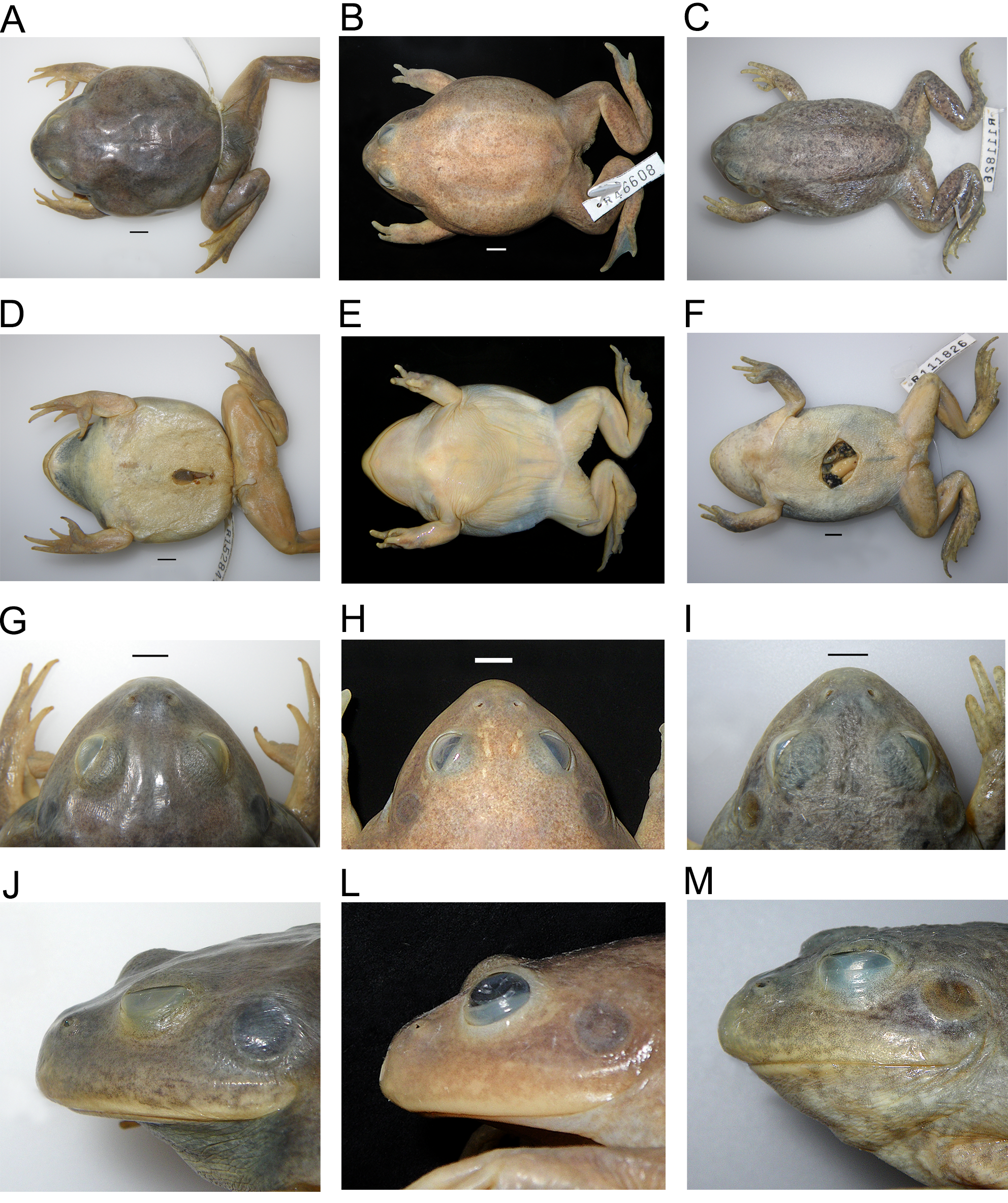
Variation. See also morphological measurements of all preserved specimens (Appendix 1; Table 2) and details of morphological variation ( Table 3 View TABLE 3 ). Head broad, somewhat depressed, and wider than long. Snout short, rounded to truncate in dorsal view and in profile, mouth wide. Eyes prominent and dorsally oriented, diameter greater than eye-to-nostril distance, with some difference noted in the northern populations (average ED/EN males/females: eastern populations = 1.36±0.18; northern 1.6±0.24). Nostrils open mainly dorsally, with slight lateral tilt, slightly closer to tip of snout than to eyes, distance between them very slightly less than eye to nostril (average IN/EN 0.92±0.12). Canthus rostralis curved, or slightly so in anterior and lateral views ( Fig. 7 View FIGURE 7 ), may be almost nonexistent in some very short-snouted northern specimens. Slight ridge from eye to nostril in some. Tympanum distinct, diameter less than eye diameter (average Tymp/ED = 0.80±0.07); annulus slightly raised, supra-tympanic skin fold distinct or indistinct and extends from edge of eye, above tympanum and around its posterior rim, terminating above forearm. Toes fully webbed, fingers unwebbed, thumb opposable. Dorsal skin mostly smooth with or without granular sides, scattered low tubercles present in some and occasionally a few short skin folds (see Table 3 View TABLE 3 ). Ventral surface smooth.
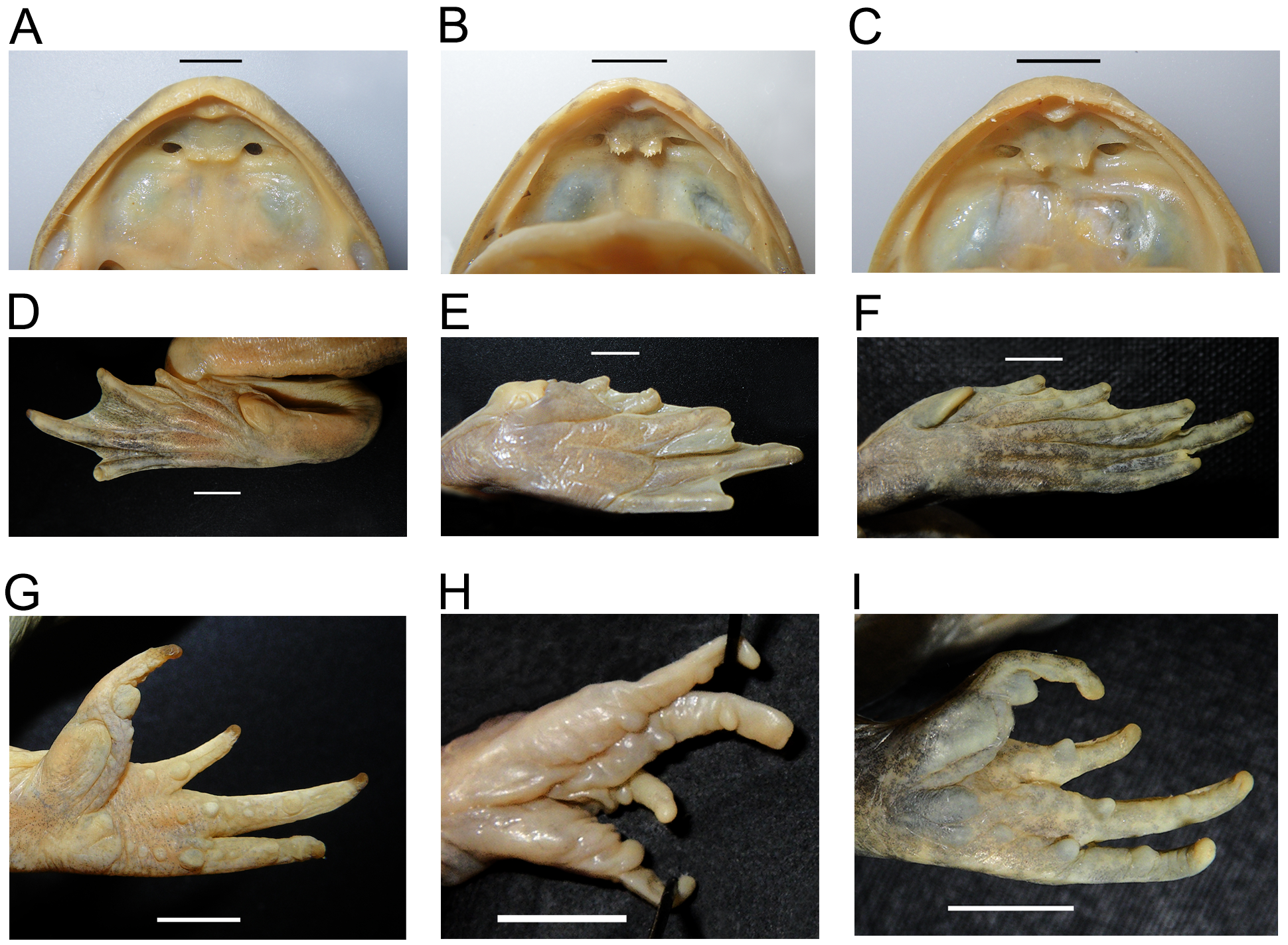
Tongue very broad, attached anteriorly, free behind. A prominent vomerine tooth plate on either side of midline, each narrowly separated and laterally adjacent to medial edge of choanae; each tilts slightly posteriorly towards medial edge; ventral edge denticulate with 5–7 small pointed tips; choana almost equal in width to vomerine tooth plate ( Fig. 8 View FIGURE 8 A,B).
Arms fairly short (FA/SVL = 0.23±0.08), hindlimbs short and robust (TibL/SVL 0.39±0.04). Fingers unwebbed, decreasing in order of length 3˃4˃1˃2; subarticular tubercles well defined, inner metacarpal tubercle large, oval and fleshy, tapering anteriorly; toes decreasing in order of 4˃5˃3˃2˃1, fully webbed to tips but tapering to broad lateral fringes towards tip of fourth toe; subarticular tubercles poorly defined and small, inner metatarsal tubercle large, ovoid and partly or fully unattached beneath medial edge; tips of fingers and toes slightly fleshy, no wider than digit and no circum-marginal grooves ( Fig. 8 View FIGURE 8 G,H).
Colour and skin texture in life. Dorsum ranges from grey, often with green areas and sometimes pinkish tinges, to dull yellow-brown (with or without darker flecks or patches) or uniform green. Specimens from the grey and brown clay soils of the Bourke/Nyngan/Gilgandra regions in western NSW usually have smooth skin, and adults tend to have a grey dorsum with green areas (with or without scattered flecks or pinkish tinges) or are sometimes all dull green. Specimens from south-western Qld are dull yellow-brown with or without reddish tinges ( Fig. 9 View FIGURE 9 ). Some specimens have a paler green or cream vertebral stripe ( Fig. 9 View FIGURE 9 G). Limbs and upper lip usually with darker flecks or mottling. Further north in the Boulia region in south-western Qld, the skin is more granular, with scattered tubercles, and frogs in life tend to be dull yellow-brown (some with dull reddish tinges). Specimens found in the heavy grey clay soil areas from the north-eastern locality east of Clermont, Qld are immaculate light grey with faint greenish tinges and smooth skin ( Meyer & Agnew 2013). Those from the Barkly Tableland, NT mostly have smooth grey skin (29% slightly granular), with a pink wash over the eyelids, along either side of the vertebral region and on the dorsal surface of the limbs ( Fig. 9 View FIGURE 9 H, K, N), and some have scattered olive-grey flecks ( Fig. 9 View FIGURE 9 B, E). Belly white; throat dark grey in calling, adult males. Iris golden.
Colour in preservative. Dorsal colour grey, grey-brown or brown, with or without faint darker flecks or mottling; some specimens (3%) with pale-grey vertebral stripe. No specimens from the northern populations have a pale vertebral stripe. Limbs and upper lip usually mottled or flecked. Belly cream or white, throat dark grey in adult males.
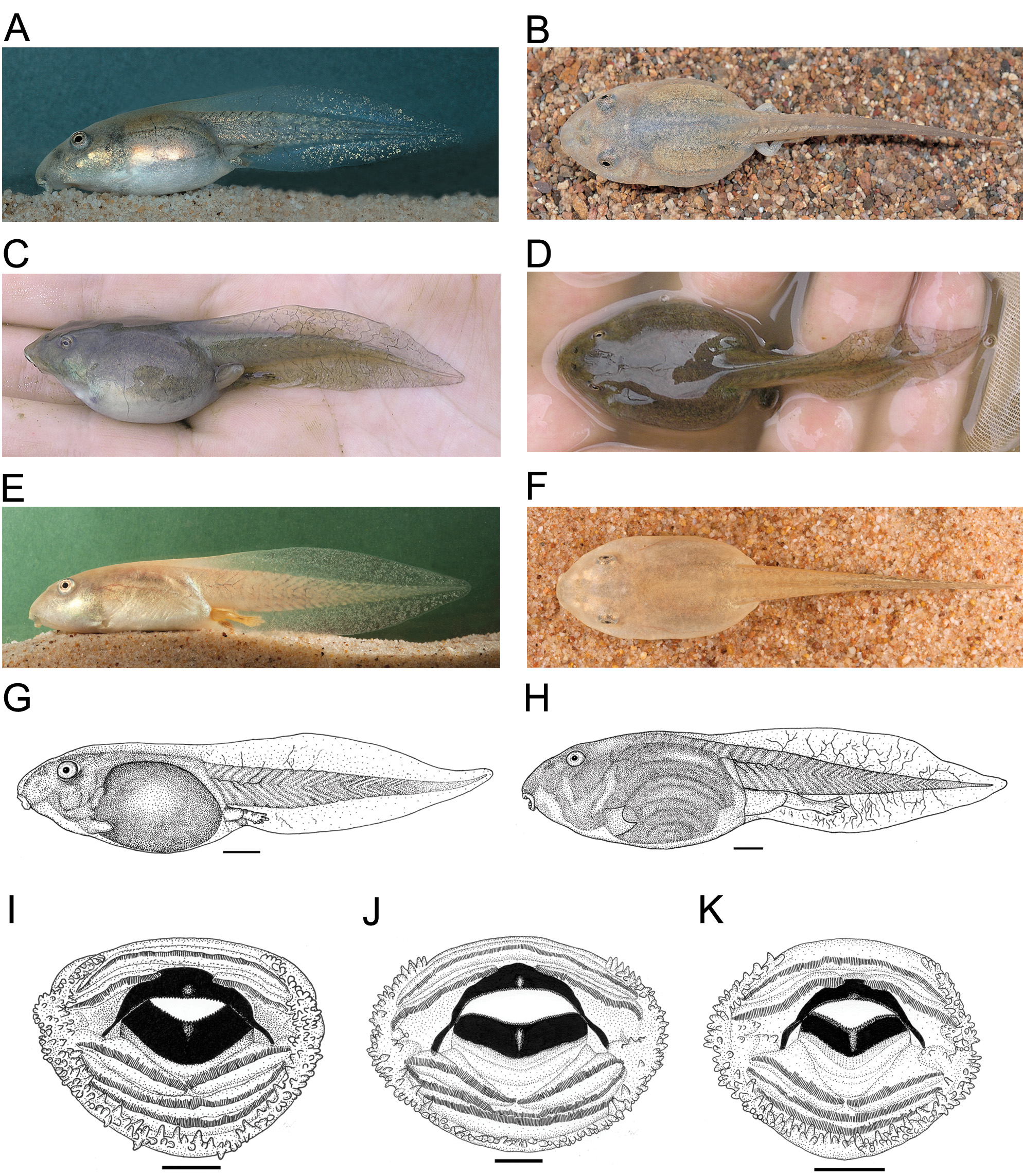
Tadpoles. Tadpoles were described by Anstis (2013). Photographs in life of a specimen from eastern and northern regions and morphometric measurements are provided here for comparison ( Fig. 10 View FIGURE 10 ; Table 4). Maximum size: TL to 109 mm, BL to 37 mm (stage 41, Bourke, NSW), but many only grow to about 70–80 mm in total length. Tadpoles of C. platycephala have moderately large to very large, plump bodies with dorsolateral eyes and moderately arched fins.
Colour in life. Tadpoles in earlier stages 25–26, as observed in the field from an eastern population, have a fine layer of gold over dorsum and may have a darker line along either side of vertebral region with a darker area over base of body. As they grow the dorsum becomes more uniformly pigmented with a layer of gold, merging with opaque silver-white and a copper-gold or greenish sheen over sides of body and venter. Iris gold. Tail muscle in fully-grown tadpoles light or darker grey-brown, with gold flecks over sides. Fins transparent to dusky dark grey, densely speckled with gold and often green iridophores. Tadpoles from northern populations on the Barkly Tableland were only observed in later stages, but were dark olive-brown over the dorsum merging with a green sheen and dull opaque silver-white over sides and on venter.
Colour in preservative. All gold and silver-white pigment is lost in preservative and tadpoles are dull and more translucent with darker internal organs visible.
Oral disc. The key characters summarised from Anstis (2013) are: broad anterior medial gap in border of marginal papillae, two anterior and three posterior tooth rows with medial gap in second anterior and first posterior rows, robust jaw sheaths, upper broadly arched.
Metamorphosis. The duration of larval life ranges from 30–50 days ( Anstis 2013), but in hotter regions in summer may be much less. Metamorphosis has been observed in late October to May in eastern populations from NSW (spring to autumn), and in late summer (February) in the Barkly Tableland, NT, but in areas of low and unpredictable rainfall, could occur at any time from spring to autumn. Metamorphs strongly resemble the adult in colour and skin texture, but the tympanum is not distinct initially. Fourteen recent metamorphs from tadpoles raised in captivity from eastern populations had a mean SVL of 27 mm (22.0–38.0 mm) and two from the Barkly Tablelands were 26.0 and 25.5 mm ( Anstis 2013). SVL of recent metamorphs observed in the field were similar to those raised in captivity, but none were measured.
Habitat. Tadpoles of C. platycephala have been observed in muddy, grey or brown water and clearer ephemeral water bodies in clay-soil areas, where they tend to be whitish-gold (muddy water) or yellow-ochre (clearer water) when fully grown, with a silver-white belly. Some have also been observed in the field with black tails at night and early morning ( Meyer & Agnew 2013; M. Anstis, unpubl. data).Those at more advanced stages from the Barkly Tableland were dull grey to olive-brown above in clearer water over black, clay substrate.
Etymology. The specific epithet is Latin for ‘flat’ (platy) and ‘headed’ (cephala).
Distribution and habitat. Occurs across a wide area of eastern Australia in lower rainfall inland areas of NSW, Qld, southern NT and northern SA ( Fig. 1 View FIGURE 1 ). There are specimens from more than 10 Interim Biogeographic Regionalisation of Australia (IBRA) bioregions ( Thackway & Cresswell 1995; Department of Environment 2012), but principally, in decreasing order of number of specimens, from: Channel Country, Mulga Lands, Darling Riverine Plains, Stony Plains, Cobar Peneplain and Riverina Bioregions (Atlas of Living Australia website www.ala.org.au, accessed 1 Sep 2015). They commonly occur in clay-based soils where water is more likely to be held for longer periods at the surface after heavy rain. Within the Brigalow Belt, for example, they avoid sandy cypress and sandstone areas in favour of adjoining communities such as brigalow, belah and coolabah on clay soil (H.B. Hines & E. Meyer, unpubl. data). After active periods, they burrow into clay soil, where they remain cocooned in their sloughed skin within a small cavity, sometimes for several years (van Beurden 1982).
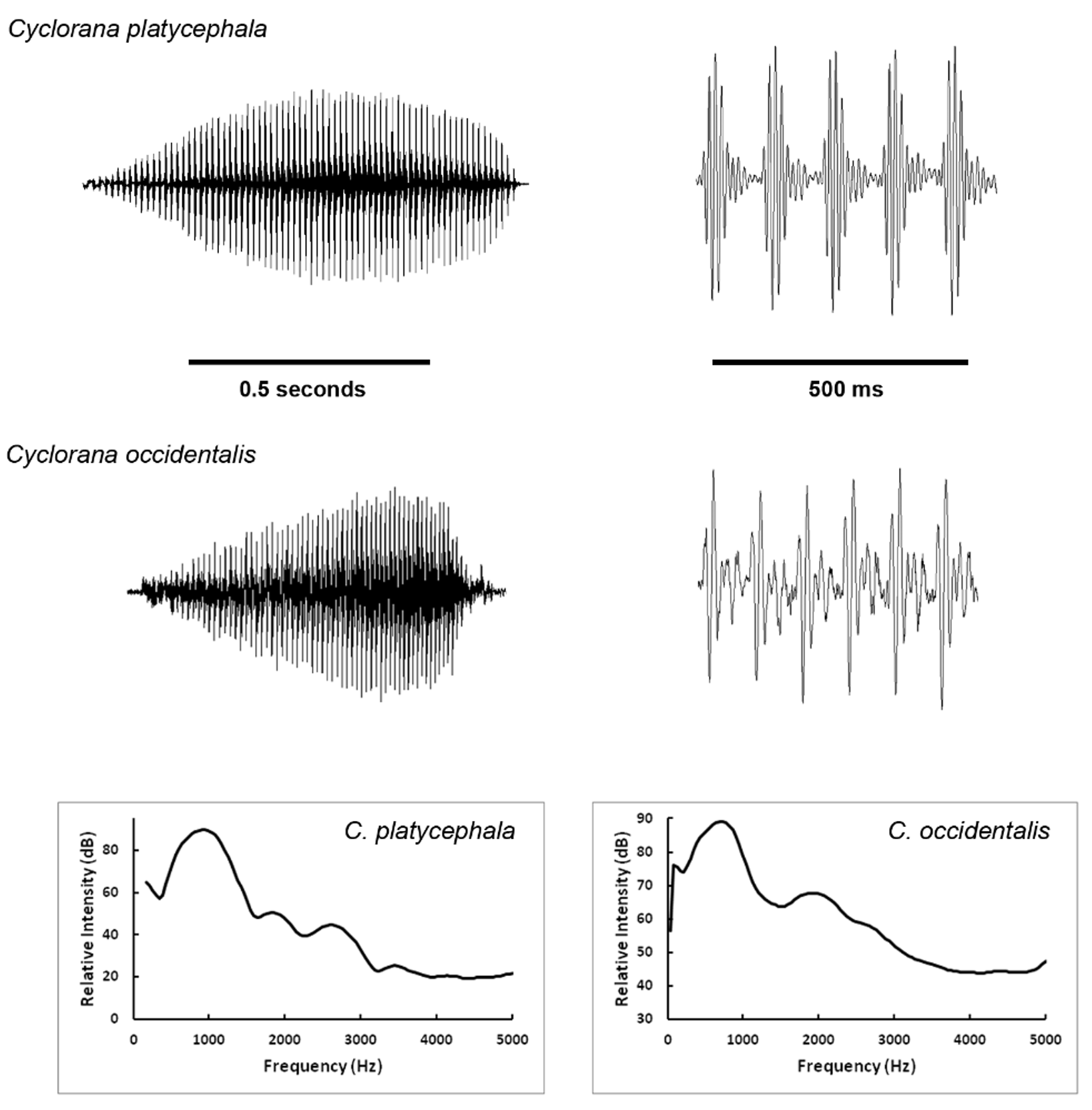
Advertisement call. The call is a long continuous pulse train, rising in amplitude mid-call, then declining relatively gradually, resulting in a nasal “ maaaaaaaah” repeated every 1–2 seconds ( Fig. 5 View FIGURE 5 ; Table 5).
Breeding behaviour. An explosive breeder, with breeding occurring in ephemeral wetlands within a few hours of heavy rainfall in warmer months. Males call while partly submerged or afloat in water. Calling, and presumably breeding, drops off considerably or ceases within a day or two of the rainfall event. In NSW, Lemckert & Mahony (2008) showed calling records for this species in October, November, March and April. In Qld, calls have been heard in November, December, February and March, with amplectant pairs observed in February (H.B. Hines, unpubl. data). Small tadpoles at stage 25 have been collected in October (M. Anstis, unpubl. data). From the detailed studies on the natural history of C. platycephala undertaken in the Coonamble-Walgett district of New South Wales (van Beurden 1982), breeding does not seem to occur following heavy rainfall in winter.



| NSW |
Royal Botanic Gardens, National Herbarium of New South Wales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chiroleptes platycephala
| Anstis, Marion, Price, Luke C., Roberts, J. Dale, Catalano, Sarah R., Hines, Harry B., Doughty, Paul & Donnellan, Stephen C. 2016 |
slevini
| Loveridge 1950 |
Chiroleptes platycephalus Günther, 1873
| Gunther 1873 |