
Pseudopothea paulai, Gil-Santana, Hélcio R., 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.3904.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:DCD221C2-DA19-4B8D-8A7B-498AF8345000 |
|
DOI |
https://doi.org/10.5281/zenodo.6094003 |
|
persistent identifier |
https://treatment.plazi.org/id/505A87A2-FF8C-FFBB-FF69-67FB61FFC0CA |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudopothea paulai |
| status |
sp. nov. |
Pseudopothea paulai View in CoL sp. nov.
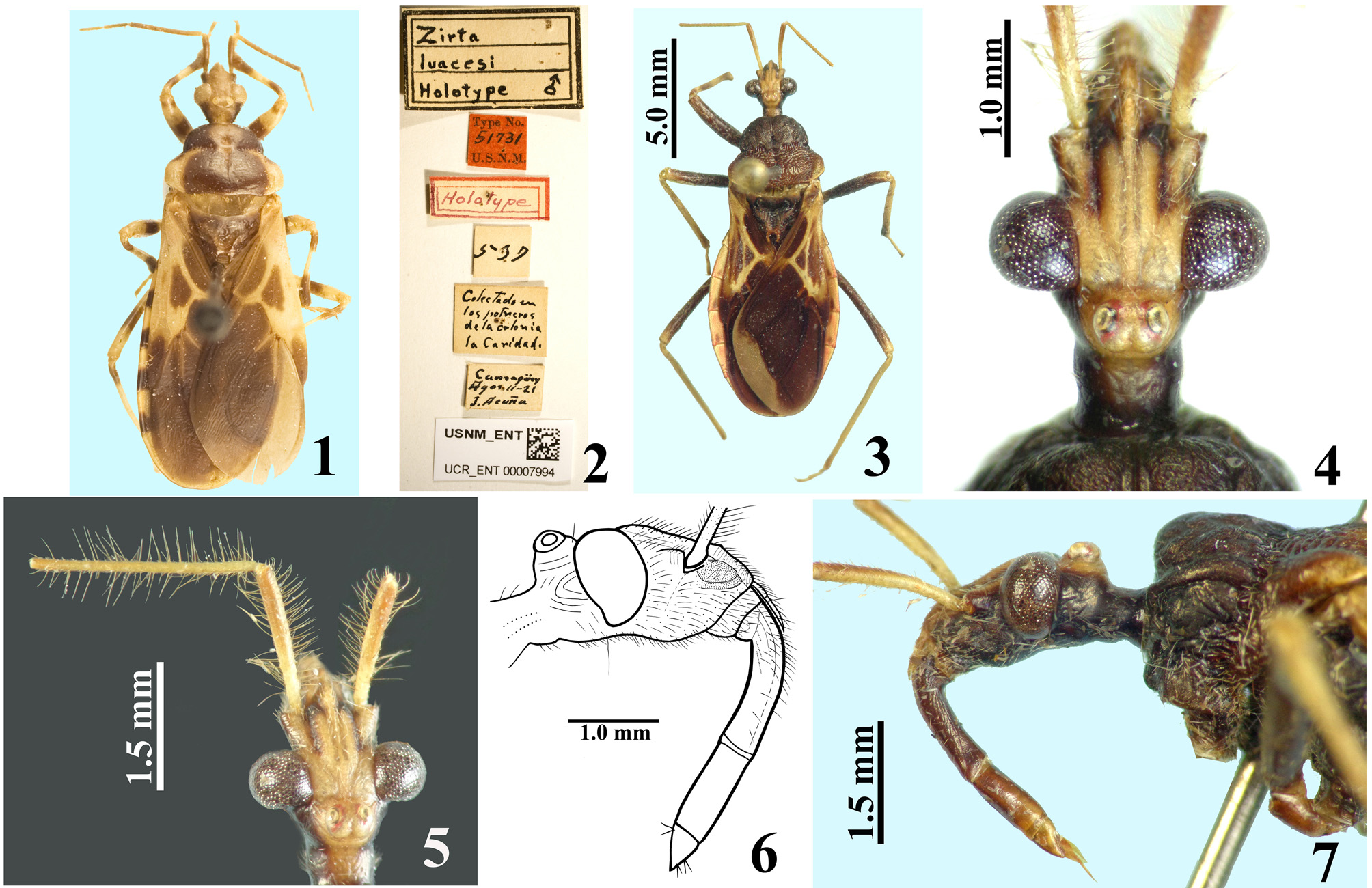
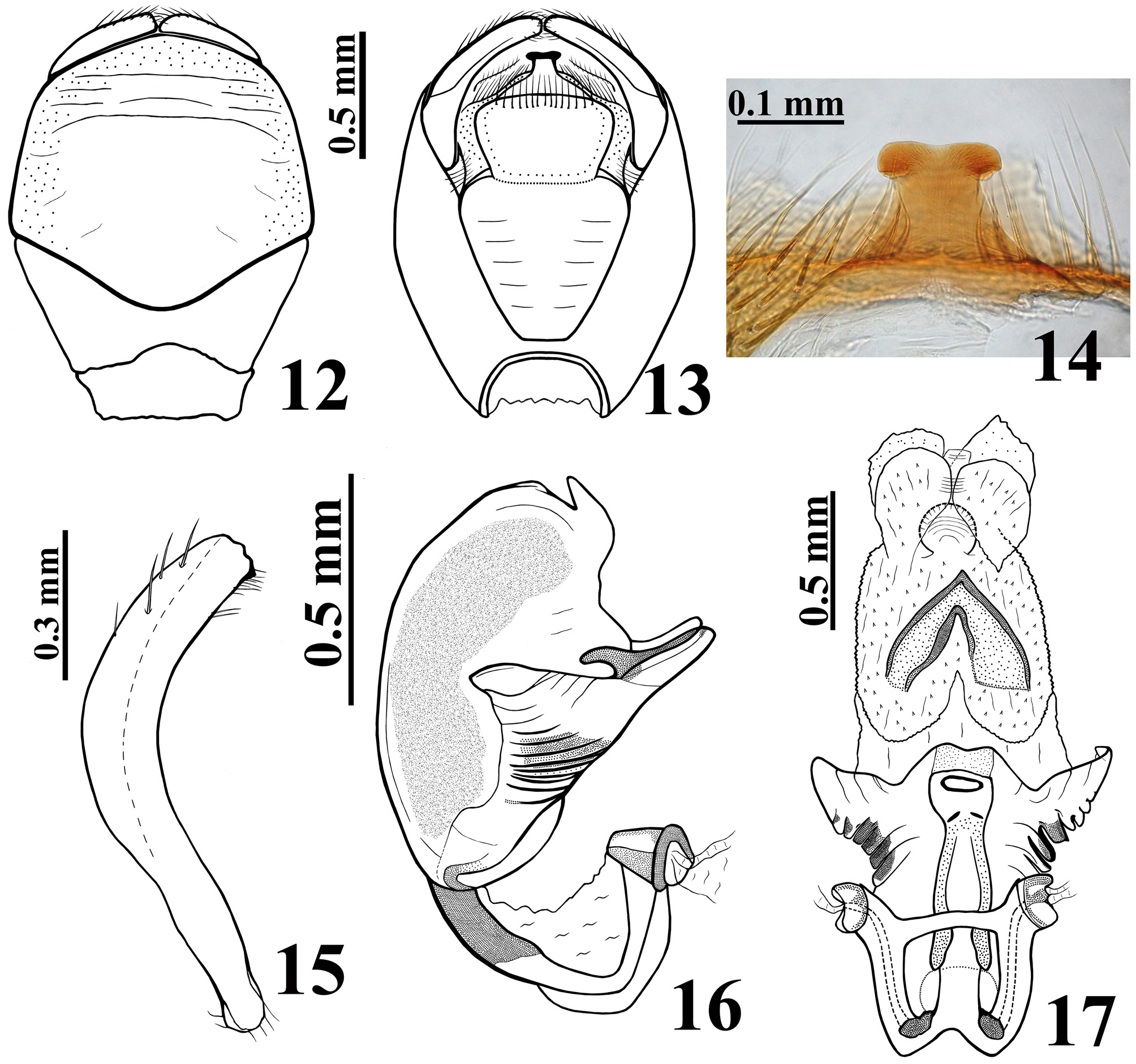
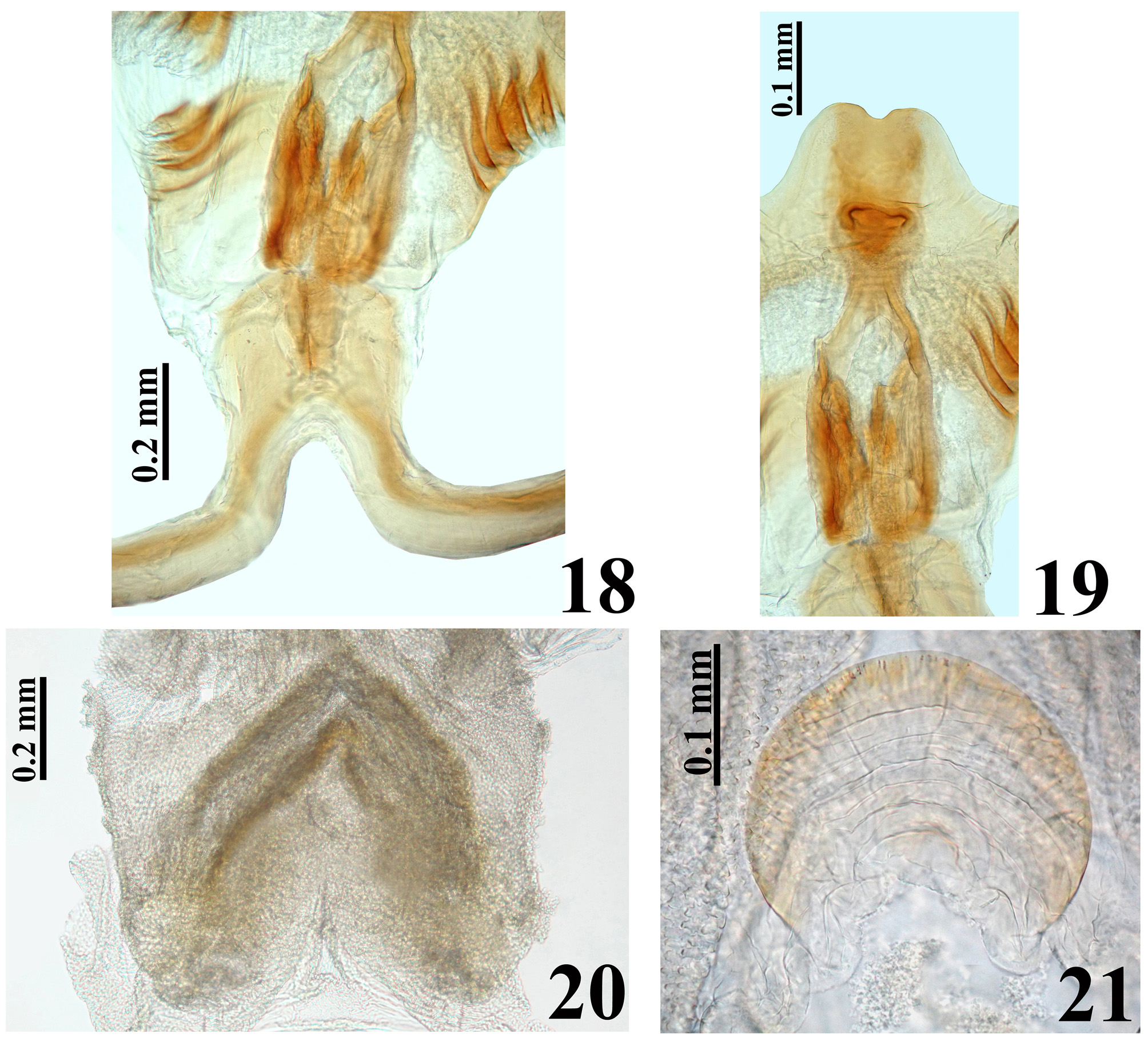
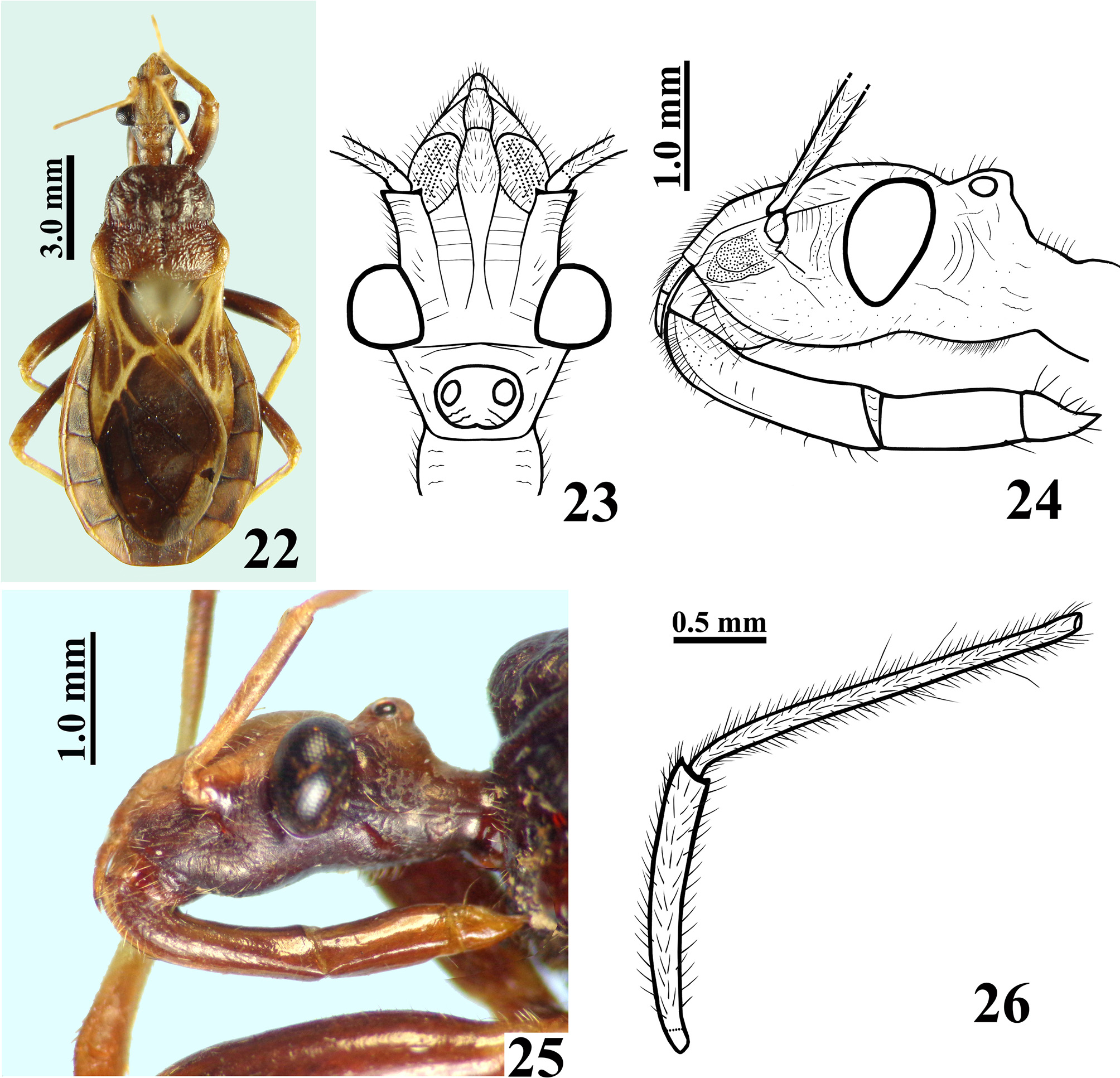
Figures 3–30 View FIGURES 1 – 7. 1 – 2 View FIGURES 8 – 11 View FIGURES 12 – 17 View FIGURES 18 – 21 View FIGURES 22 – 26 View FIGURES 27 – 30 .
Diagnosis. Pseudopothea paulai sp. nov. can readily be separated from P. luacesi ( Fig. 1 View FIGURES 1 – 7. 1 – 2 ) by the conspicuously setose and rugous integument of head and thorax, which are glabrous and smooth respectively in the latter species. The anterior half of the ocellar tubercle is at the level of the posterior margin of the eyes in P. luacesi and clearly posterior to this level in P. paulai sp. nov. The coloration is also different between these two species, mainly in terms of more extensive yellowish markings in the hind lobe of pronotum, femora and corium of hemelytra in P. luacesi . On the other hand, the grayish apex of the membrane of hemelytra and the median longitudinal yellowish wavy band on the sternites of P. paulai sp. nov. are quite distinctive. Most connexival segments are divided dorsally approximately into two halves of distinct colors: yellowish anteriorly and dark posteriorly in P. luacesi , while P. paulai sp. nov. has darkened spots of variable sizes in the posterior portion of each connexival segment.
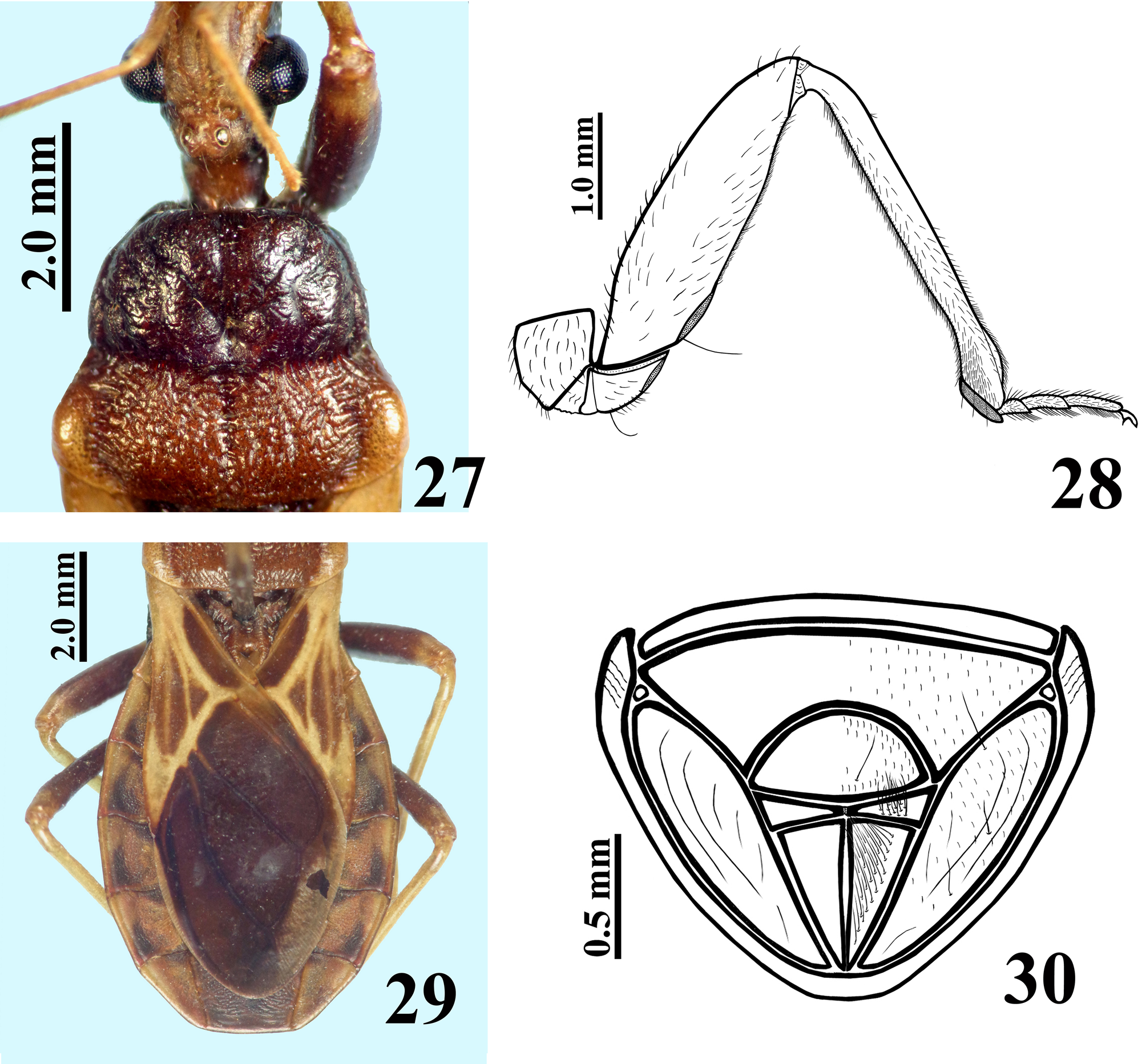
Description. Male. MEASUREMENTS: Total length: 14.5; head: total length (including collum): 2.9; (excluding collum): 2.7; maximum width across the eyes: 2.1; anteocular length: 1.2; postocular length (measured to anterior margin of collum): 0.6; interocular space: 0.8; height of head (maximum distance between upper and lower margin of the head): 1.25; right eye: transverse width: 0.65; length, measured from above: 0.7; distance between upper and lower margin: 1.0; ocellar tubercle width: 0.55; antennal segments length: I: 1.8; II: 2.7; others absent; labial segments length: II [first visible]: 1.5; III: 1.0; IV: 0.4. Thorax: pronotum: fore lobe length: 1.6; maximum width: 2.8; hind lobe: length: 1.5; width at posterior margin: 3.9; hemelytra length: 8.8. Legs length: fore legs: femur: 3.4; tibia: 3.7; spongy fossa: 0.40; tarsus: absent; middle legs: femur: 3.8; tibia: 4.0; spongy fossa: 0.35; tarsus: 1.2; hind legs: femur: 5.0; tibia: 6.1; tarsus: 1.3. Abdomen: length: 7.0; maximum width: 5.4. COLORATION: General coloration brownish with yellowish markings or portions of body ( Figs 3 View FIGURES 1 – 7. 1 – 2 , 10–11 View FIGURES 8 – 11 ). Head ( Figs 4–5, 7 View FIGURES 1 – 7. 1 – 2 ): brownish black, with antennal segments I and II (others absent) yellowish; distal two-thirds of dorsal portion and distal one-third of ventral portion of antennal segment I slightly darkened; a yellowish dorsal band that includes the median area of collum, ocellar tubercle, interocular space, antennifers and most of the area between the latter, and also the clypeus and labrum and areas adjacent to these two; labium brownish with distal portion of segment III and segment IV somewhat clearer; a thin reddish stripe around ocelli present; eyes black. Thorax ( Figs 3, 7–8, 10 View FIGURES 1 – 7. 1 – 2 View FIGURES 8 – 11 ): hind lobe of pronotum slightly clearer with humeral angles somewhat yellowish; pleural and sterna area darkened, brownish black. Legs: coxae and trochanters brownish; femora brownish with a small portion of apex, dorsally, clearer to yellowish. Tibiae yellowish; fore tibia with base, a subbasal large ring and apical fifth brownish; middle and hind tibiae somewhat darker at apex and with small faint basal and subbasal darkened markings. Tarsi yellowish with apex of third segment somewhat darkened. Hemelytra fuscous, corium mostly yellowish, with the area adjacent to scutellum, apex and two or three ill-defined stripes in the central portion, brownish; membrane brownish, basal portion of the veins somewhat brighter, with almost all the apical portion of membrane, external to and bordering the cells, grayish. Abdomen ( Figs 10–11 View FIGURES 8 – 11 ): connexivum dorsally yellowish with a light reddish tint and faint small darkened spots on middle of distal portion of visible segments, which do not reach the intersegmental suture; ventrally, yellowish. Sternites brownish with portions of sternites adjacent to connexivum yellowish too, forming a lateral wavy band, enlarged in each sternite in the area of the spiracle, which is included in the yellowish portion of this band; sternites III–VII with a median longitudinal wavy yellowish band; on the sternite VII, the yellowish band ends a little above the posterior margin of the segment; pygophore brownish. VESTITURE: Head: covered with slender elongate setae, more numerous on clypeus and labrum, and also on adjacent portions ( Figs 4–7 View FIGURES 1 – 7. 1 – 2 ); antennal segments I and II (others absent) covered with long yellowish thin erect hairs of 0.35 to 0.45 mm in length, except on dorsal portion of first segment where numerous shorter yellowish setae of approximately 0.1 mm in length are present ( Fig. 5 View FIGURES 1 – 7. 1 – 2 ); labium with yellowish elongate setae somewhat numerous on ventral and lateral faces of labial segment II; one pair on base, dorsally, and a few on apex of labial segment IV ( Fig. 6 View FIGURES 1 – 7. 1 – 2 ). Thorax: yellowish to whitish slender setae very sparse on pronotum, to longer and considerably numerous setae on sterna. Legs covered with numerous moderate long yellowish setae, which are even more numerous on apices of tibiae ( Fig. 9 View FIGURES 8 – 11 ); elongate narrow patches of stiff short hairs on ventral face of fore and middle trochanters and femora, on these latter in basal portion ( Fig. 9 View FIGURES 8 – 11 ); fore and middle femora and tibiae with a ventral fringe of short curved setae ( Fig. 9 View FIGURES 8 – 11 ); apex of tibiae and tarsi covered with somewhat longer setae with same coloration. Scutellum and hemelytra glabrous. Abdomen: connexivum glabrous; sternite II with long hairs on median portion; other sternites with very small and sparse adpressed setae, mainly on median portions of the segments. STRUCTURE: Head ( Figs 4–7 View FIGURES 1 – 7. 1 – 2 ): elongate, length more than twice the height of head (maximum distance between upper and lower margin of the head); anteocular portion about twice as long as postocular; integument rugous, with linear irregular impressions, smooth on labium segments III and IV; tubercle ocellar undivided; transverse sulcus located just behind eyes, shallow; eyes salient, globose in dorsal view, suboval in lateral view, reaching dorsal margin but not ventral margin of the head; clypeus slightly pointed in lateral view; large antennifers; first antennal segment stout, somewhat curved, base slender, the latter of approximately 0.15 mm in width, whereas on middle to apical third the width is approximately 0.21 mm; second segment slender, cylindrical, of 0.12 mm in width; labium thickened, labial segment II (first visible) very little longer than III and IV together, reaching or slightly surpassing level of anterior margin of eyes in lateral view. Thorax: anterolateral angles rounded and small; fore lobe approximately same length as and narrower than hind lobe of pronotum ( Fig. 8 View FIGURES 8 – 11 ); fore lobe rounded on anterior and lateral margins; a somewhat large but shallow lateral ridge on lateral portion ( Fig. 7 View FIGURES 1 – 7. 1 – 2 ); integument diffusely rugous on fore lobe and transversally rugous on hind lobe of pronotum ( Fig. 8 View FIGURES 8 – 11 ); median longitudinal sulcus very thin and shallow, with a pair of parallel lines beside it, which run in anterior twothirds of the fore lobe, ending in a moderately deep median fovea, which is separated from the transverse (interlobar) sulcus by a thin, curved carina; the transverse sulcus is formed by small and shallow impressions (canaliculae), interrupted medially; on hind lobe of pronotum the median longitudinal sulcus is somewhat deeper, ending approximately at distal third, far from posterior margin of hind lobe; lateral sulcus of posterior lobe of pronotum very weak, formed by strengthening of the striation of the integument; humeral angles rounded ( Fig. 8 View FIGURES 8 – 11 ). Pleural and sternal areas rugous and coarsely striated. Stridulitrum long, surpassing fore coxae and reaching mesosternum, larger on first anterior half. Fore acetabulae somewhat prominent. Metasternum elevated on central portion. Scutellum strongly narrowed posteriorly, sculptured; median depression deep with transverse small ridges, prongs thin and convergent. Fore and middle femora somewhat thicker than hind ones, the former more than the latter; width and width/length ratio of fore and middle femora respectively: 0.67 mm / 0.19; 0.60 mm / 0.15. Presence of a median ventral thin and shallow crest with very small tubercles, where the ventral fringe of short curved setae are inserted, on fore and middle femora and tibiae. Hind femora slightly thicker subapically. Fore tibia thicker at apex, with a distal comb on anterior face; middle tibiae very slightly thicker at apex. Spongy fossae very small, with about one-ninth (fore tibia) to one-eleventh (middle tibia) of respective tibial length. Hemelytra reaching tip of abdomen. Abdomen: connexivum with posterolateral angle between segments II and III somewhat prominent; sternite II striated laterally, other sternites with smooth integument and very fine striations anteriorly and laterally, somewhat more marked around genitalia on sternite VII. Sternites II and III separated by canaliculae. Spiracles elliptical. Male genitalia ( Figs 12–21 View FIGURES 12 – 17 View FIGURES 18 – 21 ): pygophore sub-squared, posterior margin sinuate; parameres apices close in resting position ( Fig. 12–13 View FIGURES 12 – 17 ). Median process of pygophore sclerotized, enlarged at apex, somewhat similar to a “T” shape ( Fig. 14 View FIGURES 12 – 17 ). Parameres symmetrical, a little curved, with a short subapical blunt tooth and a small rounded apical protuberance; sparse setae on apical half of external face and shorter and denser ones on apical portion of internal face, particularly near to subapical tooth ( Fig. 15 View FIGURES 12 – 17 ). Phallus with articulatory apparatus moderately long, subquadrate ( Figs 16–18 View FIGURES 12 – 17 View FIGURES 18 – 21 ). Dorsal phallothecal plate enlarged to the apex, sinuous in center of anterior margin and more pronouncedly sinuate laterally to anterior margin ( Figs 17 View FIGURES 12 – 17 , 19 View FIGURES 18 – 21 ); midlateral portions with pronounced grooves ( Figs 16–17 View FIGURES 12 – 17 ); struts enlarged in basal half and united at the apex, which is enlarged too with a more sclerotized large central portion ( Figs 18–19 View FIGURES 18 – 21 ). Endosoma with a pair of flat subapical lobes, a subbasal large subtriangular spiny process, and a subhemispherical somewhat sclerotized median process ( Figs 17 View FIGURES 12 – 17 , 20–21 View FIGURES 18 – 21 ).This latter has concentric thin striations ( Fig. 21 View FIGURES 18 – 21 ). Female: MEASUREMENTS: Total length: 13.8; head: total length (including collum): 3.0; (excluding collum): 2.8; maximum width across the eyes: 2.0; anteocular length: 1.5; postocular length (measured to anterior margin of collum): 0.5; interocular space: 0.9; height of head (maximum distance between upper and lower margin of the head): 1.3; right eye: transverse width: 0.5; length, measured from above: 0.7; distance between upper and lower margin: 0.9; ocellar tubercle width: 0.5; antennal segments length: I: 1.6; II: 2.0; others absent; labial segments: II [first visible]: 1.6; III: 1.0; IV: 0.5. Thorax: pronotum: fore lobe length: 1.7; maximum width: 2.9; hind lobe: length: 1.4; width at posterior margin: 3.7; hemelytra length: 7.6. Legs length: fore legs: femur: 3.3; tibia: 3.5; spongy fossa: 0.45; tarsus: 1.0; middle legs: femur: 3.8; tibia: 3.8; spongy fossa: 0.40; tarsus: 1.2; hind legs: femur: 5.0; tibia: 5.6; tarsus: 1.4. Abdomen: length: 6.5; maximum width: 5.5. COLORATION: similar to male ( Figs 22, 25 View FIGURES 22 – 26 , 27, 29 View FIGURES 27 – 30 ). Legs with somewhat more extensive yellowish markings on: fore trochanter externally; base and apex of fore femora; middle and hind trochanter and base of middle and hind femora. Corium of hemelytra with apical portion yellowish ( Fig. 29 View FIGURES 27 – 30 ). Connexivum much more darkened than in male, with larger apical brownish-black markings reaching the intersegmental suture ( Figs 22 View FIGURES 22 – 26 , 29 View FIGURES 27 – 30 ); yellowish ventrally, with posterior half of segments III–VI darkened; adjacent portion of sternites with a much narrower lateral wavy bright band, somewhat enlarged in each sternite in the area of the spiracle, which is partially included in the yellowish portion of this band; median longitudinal wavy yellowish band on sternites fainter, extending only to anterior portion of segment VII. VESTITURE AND STRUCTURE: Integument less setose than in male ( Figs 22–25 View FIGURES 22 – 26 , 28 View FIGURES 27 – 30 ). Head: eyes somewhat smaller, less salient than in male, not reaching dorsal or ventral margins of the head; anteocular portion about three times as long as postocular; rounded clypeus in lateral view ( Figs 23–25 View FIGURES 22 – 26 ); antenna (segments I and II, others absent) covered with short yellowish setae of approximately 0.1 mm in length; on segment II, a few scattered long yellowish hairs of approximately 0.25 to 0.4 mm in length ( Fig. 26 View FIGURES 22 – 26 ). First antennal segment somewhat thinner in basal third, of approximately 0.18 mm in width, whereas in middle to apical third, the width is approximately 0.21 mm; second segment thinner, cylindrical, of width 0.12 mm ( Fig. 26 View FIGURES 22 – 26 ). Thorax: fore lobe of pronotum a little longer than hind lobe. Setae less numerous on legs, particularly on coxae, trochanters and femora ( Fig. 28 View FIGURES 27 – 30 ). Fore and middle femora thicker than in male ( Fig. 28 View FIGURES 27 – 30 ); width and width/ length ratio of fore and middle femora: 0.8 mm / 0.25; 0.7 mm / 0.19, respectively. Spongy fossae very small, of about one-eighth (fore tibia) to one-tenth (middle tibia) of respective tibiae length. Hemelytra not reaching tip of abdomen, ending approximately at half of seventh tergite ( Fig. 29 View FIGURES 27 – 30 ). Female genitalia: Posterior view of external genitalia as in Fig. 30 View FIGURES 27 – 30 .
Speciments examined. Holotype: BRAZIL, Amazonas, Benjamin Constant, IX.1977, I. C. Lima [ leg.], [MNRJ], 1 male. Paratype: BRAZIL, Ceará, Carquejo, III.1963, Dirings[hofen, collection], 1 female [MZSP].
Distribution. Brazil, states of Amazonas and Ceará.
Etymology. The new species is named in honor of the Brazilian entomologist, Alexandre Silva de Paula, for his great contribution to studies on Heteroptera.
Discussion. Many of the differences between the male and female specimens of P. paulai sp. nov. examined here are in accordance with the sexual dimorphic features recorded in several other species of Ectrichodiinae , as follows. First two antennal segments (others absent) pubescent in male ( Fig. 5 View FIGURES 1 – 7. 1 – 2 ), while in female these segments have short setae ( Fig. 26 View FIGURES 22 – 26 ); eyes somewhat larger in male ( Figs 4, 6–7 View FIGURES 1 – 7. 1 – 2 , 23–25 View FIGURES 22 – 26 ); clypeus slightly pointed in lateral view in male ( Fig. 6 View FIGURES 1 – 7. 1 – 2 ); thicker fore and middle femora ( Figs 9 View FIGURES 8 – 11 , 28 View FIGURES 27 – 30 ) and shorter hemelytra in female ( Figs 3 View FIGURES 1 – 7. 1 – 2 , 10 View FIGURES 8 – 11 , 22 View FIGURES 22 – 26 , 29 View FIGURES 27 – 30 ). On the other hand, the differences observed in the coloration, the integument less setose in female and the small differences in the measurements of length and some structures as described above may be due to interindividual variation and not to sexual dimorphism. Gil-Santana (2014), for example, recorded substantial variation in color, vestiture and robustness among specimens of Pothea jaguaris ( Carpintero, 1980) , regardless of their sex. Therefore, only future examination of more specimens of P. paulai sp. nov. will make it possible to ascertain to what extent these features vary among individuals and whether there is any additional sexual dimorphism.
The inclusion of P. paulai sp. nov. in Pseudopothea is in accordance with most of the main characteristics assigned to this genus by Wygodzinsky (1951), Carpintero (1978), Dougherty (1995), Maldonado & Carpintero (1996) and Forero (2004), as stated in the new diagnosis proposed here. However, this new diagnosis does not include the number of antennal segments because this characteristic could not be verified in the specimens of P. paulai sp. nov. examined here. The distal segments of the antennae were missing in both of them. Although P. luacesi has an eight-segment antenna ( Bruner & Barber 1937) and consequently subsequent authors took this number of segments into consideration in the diagnosis of Pseudopothea ( Dougherty 1995, Maldonado & Carpintero 1996), the number of antennal segments has been shown to vary among ectrichodiines, including those belonging to the same genus, as in Brontostoma spp. ( Maldonado & Carpintero 1996). Therefore, this characteristic will only be included if or when more specimens of P. paulai sp. nov. with complete antennae are found.
There is no doubt that the main characteristics attributed to Pseudopothea by Wygodzinsky (1951) when he created the genus, i.e. a moderately thick labium, and the second [first visible] segment surpassing the level of the anterior margin of the eyes, were shown by P. paulai sp. nov. ( Figs 6–7 View FIGURES 1 – 7. 1 – 2 , 24–25 View FIGURES 22 – 26 ).
However, some characteristics that were considered diagnostic for Pseudopothea by other authors were shown to be useful for separating the new species from P. luacesi .
The smooth body surface with no pubescence, which was taken into consideration in the diagnosis of Pseudopothea by Dougherty (1995) contrasts with the conspicuously setose and rugous integument of the head and thorax of P. paulai sp. nov. Although these latter characteristics are less common in the members of Ectrichodiinae , given that they occur sporadically in several species belonging to different genera within the subfamily, consideration of these characteristics at specific and not generic level is in accordance with this pattern of occurrence among Ectrichodiinae .
The anterior half of the ocellar tubercle at the level of the posterior margin of the eyes in Pseudopothea (as observed in P. luacesi ) was considered to be one of the characteristics that separated this genus from Pothea by Carpintero & Maldonado (1996). However, quick look at on some specimens of this latter genus reveals that the same characteristic is present in Pothea furtadoi , for example. This species was recently redescribed by Gil-Santana (2014) (see his figures 29 and 30). Thus, this characteristic is of no value for the generic diagnosis, but may be helpful for separating species, as in the present case.
Although the female of P. l ua c es i was originally described as being almost completely apterous ( Bruner & Fracker 1926), while the female of P. paulai sp. nov. examined here had hemelytra that were only slightly shortened ( Figs 22 View FIGURES 22 – 26 , 29 View FIGURES 27 – 30 ), this difference was not taken into consideration for separating these species from each other. Since shortening of the wings in females of Ectrichodiinae may vary within the same species, there is a need to examine more specimens of both species in order to be sure about the possible differences in this feature between Pseudopothea spp., or the extent of such differences.
On the other hand, the opinion of Wygodzinsky (1951) that Pothea would include only very delicate species, in contrast with a more robust appearance among Pseudopothea deserves some comments. At first sight, P. paulai sp. nov. would seem to be just as robust as P. luacesi ( Figs 1, 3 View FIGURES 1 – 7. 1 – 2 , 22 View FIGURES 22 – 26 ), which would be in accordance with its inclusion in Pseudopothea, following Wygodzinsky’s (1951) generic concept. However, as emphasized by Carpintero (1978), while some species of Pothea are graceful and delicate, others are clearly robust, a fact which has been recorded by other authors ( Carpintero 1980, Maldonado & Carpintero 1996, Gil-Santana 2014). Therefore, this characteristic is not useful for separating these two genera and, as this is a common feature in many other Ectrichodiinae in several genera, it is not recommended as a diagnostic feature of Pseudopothea, either.
Although some of the recorded sexual dimorphic features found in Ectrichodiinae in general were recorded in P. paulai sp. nov., as discussed above, extreme brachyptery, broader abdomen, larger head, smaller ocelli, posterior pronotal lobe and scutellum, as recorded in relation to females of P. luacesi ( Bruner & Fracker 1926, Dougherty 1995), were not confirmed in the female of P. paulai sp. nov., in comparison with the male examined here. As discussed earlier, only future examination of more specimens of both species of Pseudopothea will make it possible to ascertain the extent to which these features vary among individuals and, therefore, whether they can or cannot be considered to be sexually dimorphic features.
It is noteworthy that Ectrichodiella, which just like Pseudopothea, was previously known only in Cuba and is considered to be more closely related to Old World genera of Ectrichodiinae ( Wygodzinsky 1951) , was recently also recorded in the Amazonian region of Brazil ( Gil-Santana et al. 2013). These new records of Neotropical Ectrichodiinae show that these taxa, which previously had been considered isolated or vicariant, have a hitherto unsuspected geographical distribution.
Recently, Gil-Santana (2014) described the male genitalia of three species of Pothea , in which several differences were recorded, mainly with regard to the shapes of the parameres, the median process of the pygophore, the dorsal phallothecal plate and struts and the median process of the endosoma, which appear to be useful in relation to the taxonomy of this group. The dorsal phallothecal plate and struts were considered particularly noteworthy, since their shape and “design” are invariable within species and seem very particular to each species. In P. paulai sp. nov., these structures also seem to be very conspicuous, and the description and figures furnished here ( Figs 14–19, 21 View FIGURES 12 – 17 View FIGURES 18 – 21 ) could be useful in future taxonomic studies dealing with other species, thereby helping to clarify the systematics of the group.
On the other hand, some features of the male genitalia of P. paulai sp. nov. are very similar to those of Jorgcoris aczeli as can be judged by the figures furnished by Wygodzinsky (1959) relating to this latter species, including the parameres and particularly, the median process of the pygophore and the dorsal phallothecal plate. These two species also have in common the peculiar clear stripe bordering the cells in the apical portion of the membrane of hemelytra ( Figs 3 View FIGURES 1 – 7. 1 – 2 , 10 View FIGURES 8 – 11 , 22 View FIGURES 22 – 26 , 29 View FIGURES 27 – 30 ).
The similarities observed among the species of Pseudopothea and those belonging to genera considered close to it, such as Pothea and Jorgcoris, need to be better evaluated through more extensive taxonomical studies. Future cladistic analyses should be carried out to assess the taxonomic validity of genera and their systematic positions, and the relationships among species of New World genera of Ectrichodiinae .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ectrichodiinae |
|
Genus |