
Metacyclops thailandicus, Boonyanusith & Sanoamuang & Brancelj, 2018
|
publication ID |
https://doi.org/10.5852/ejt.2018.431 |
|
DOI |
https://doi.org/10.5281/zenodo.5640772 |
|
persistent identifier |
https://treatment.plazi.org/id/4D0287B1-FF9E-531B-FDBB-199C2F150144 |
|
treatment provided by |
Plazi |
|
scientific name |
Metacyclops thailandicus |
| status |
sp. nov. |
Metacyclops thailandicus sp. nov.
urn:lsid:zoobank.org:act:5AA8A750-7102-4258-9992-52DB66BDE4F0
Figs 6–8 View Fig. 6 View Fig. 7 View Fig. 8
Etymology
The specific epithet is derived from the country ( Thailand) where the species was collected. It is a noun in the nominative singular masculine.
Type material
Holotype
THAILAND: ♀, 475 µm long, Ratchaburi Province, Khao Bin Cave , 13°35′25.6″ N 99°40′13.1″ E, 78 m a.s.l., filtering bottle, 21 Nov. 2009, C. Boonyanusith leg., completely dissected and mounted on a slide in glycerol and sealed with nail polish ( NHM 2011.2085 About NHM ). GoogleMaps
Paratypes
THAILAND: 2 ♀♀ without egg sacs, 1 ♀ with egg sacs, sampled with the holotype, whole specimens stored in 70% alcohol (NHM 2011.2086‒2088 ) GoogleMaps ; 2 ♀♀ without egg sacs, same data as holotype,whole specimens stored in 70% alcohol ( KKU-COP-2011-004 ) GoogleMaps ; 2 ♀♀, 2 ♀♀ with egg sacs, same data as holotype, whole specimens stored in 70% alcohol (NIB). GoogleMaps
Type locality
The Khao Bin Cave is situated in the Muang District, about 20 km west of the town of Ratchaburi. The cave is in a small limestone valley located in a national forest reserve area. It is an about 300 m long fossil cave, with horizontal galleries only. The entrance is at an elevation of 78 m a.s.l. The coordinates of the entrance are 13º35′25.6″ N, 99°40′13.1″ E. Beyond the entrance, which is at the level of the valley’s floor, is a horizontal gallery, easily accessible. The gallery is about 10 m wide and 8‒10 m high, well decorated with stalagmites and stalactites. Some temporary pools are present, filled by dripping water in the rainy season. In the innermost part, there is a permanent pool, with a diameter of about 1 m and a depth of about 50 cm, filled mainly by seeping water and occasionally by dripping water. This is the type locality of Metacyclops thailandicus sp. nov. On 21 Nov. 2009, the water depth was 45 cm; the water was transparent, with a temperature of 25.1°C, pH 8.15 and conductivity of 557 µS cm-1. There was a slight amount of fine organic debris in the pool.
Description
Female
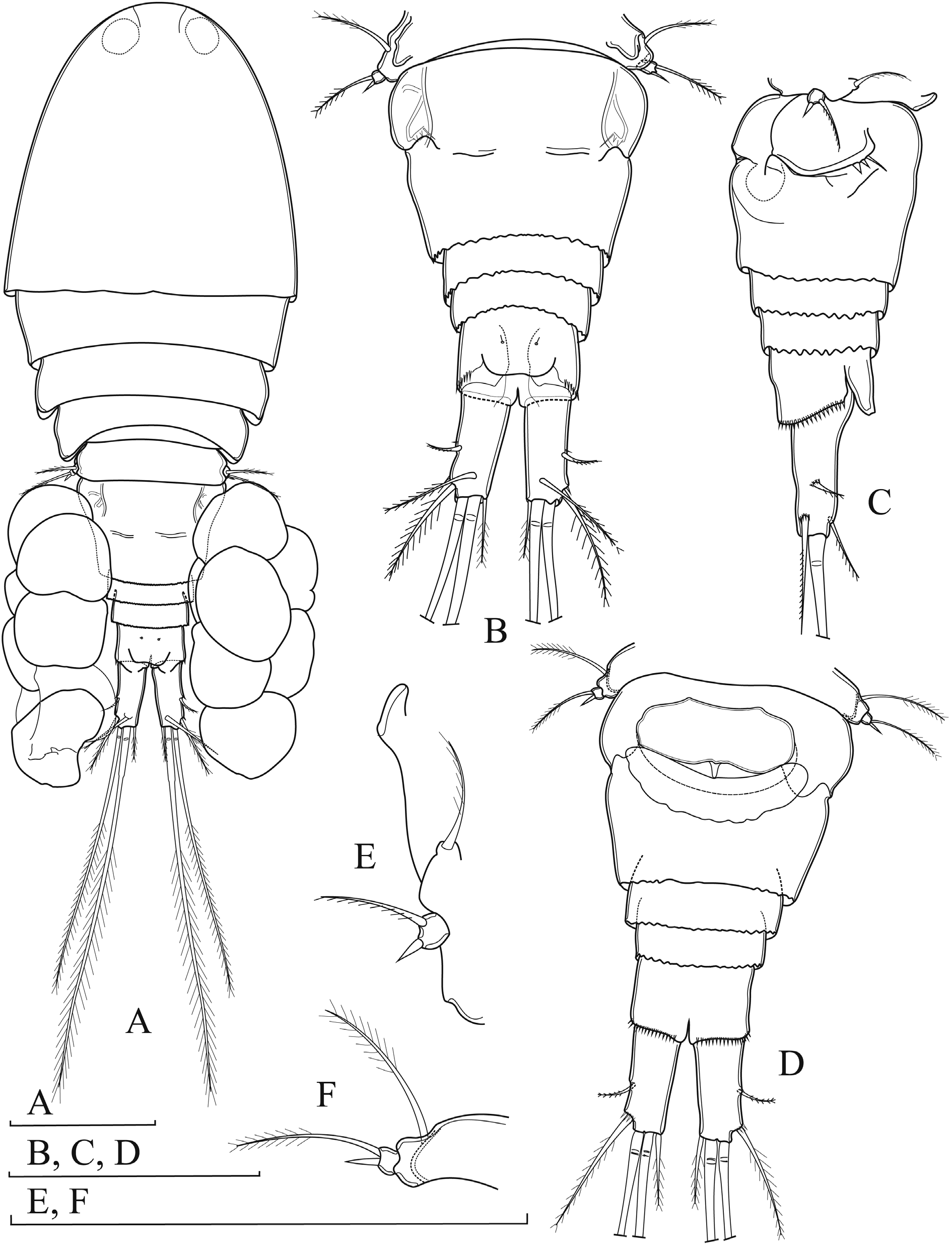
Body length, measured from tip of rostrum to posterior margin of caudal rami, 443‒486 µm (mean: 473 µm; n = 10); prosome/urosome ratio about 2.2 ( Fig. 6A View Fig. 6 ). Body dorsoventrally depressed. Preserved specimens transparent; naupliar eye not discernible; rostrum small, triangular. Cephalothorax anteriorly oval, as long as wide, representing 40 % of body length; length/width ratio about 1.1, with greatest width at posterior end. Posterior margins of Pd1–Pd4 smooth. Integument smooth, not strongly chitinized, no cuticular windows visible. Body length/width ratio about 2.5; cephalothorax about 1.8 times as wide as genital double-somite. Genital double-somite well developed, about 0.7 times as long as rest of urosome, including caudal rami; tapering posteriorly and slightly indented at level of seminal receptacle ( Fig. 6B, D View Fig. 6 ). Genital double-somite about 1.25 times as wide as long ( Fig. 6B, D View Fig. 6 ). Copulatory pore small, ovoid, situated near ½ length of double-somite; copulatory duct short, narrow and well sclerotized. Seminal receptacle with clear separation between anterior and posterior expansions; both expansions short; posterior more sclerotized and expanded laterally ( Fig. 6D View Fig. 6 ). Ovipores situated laterally, covered with reduced P6 ( Fig. 6C View Fig. 6 ). Posterior border of genital double-somite and two subsequent urosomites ornamented with hyaline fringes with irregular indentation dorsally and lateroventrally; slightly undulated ventrally. Anal somite with anal operculum reaching articulation of caudal rami; free margin smooth, slightly concave; two large sensilla at base of anal operculum ( Fig. 6B View Fig. 6 ).
CAUDAL RAMI ( Fig. 6B–D View Fig. 6 ). Slightly divergent; each ramus about 2.6 times as long as wide. Anterolateral accessory seta (I) reduced. Anterolateral seta (II) bipinnate, implanted at about ⅔ length of ramus, shorter than ramus, near implantation of seta ornamented with few spinules. Posterolateral seta (III) slim, plumose, as long as ramus. Outer and inner apical setae (IV, V) plumose; with fracture planes; inner apical seta (V) 1.3 times as long as outer one (IV) and about 0.5 times as long as body length. Apical accessory seta (VI) straight, about 0.5 times as long as ramus. Dorsal seta (VII) plumose, inserted at about ¾ length of ramus; as long as ramus.
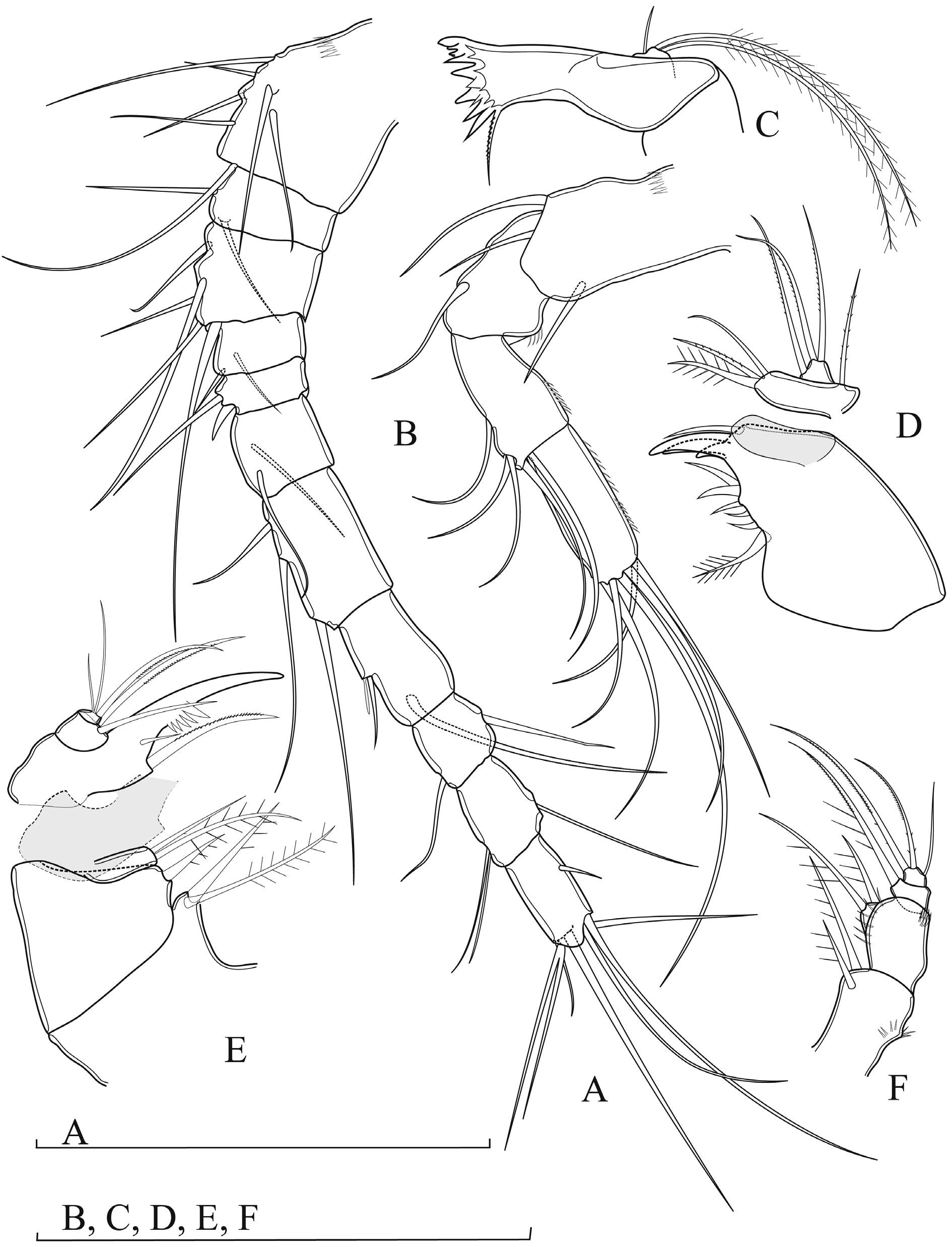
ANTENNULE ( Fig. 7A View Fig. 7 ). 11-segmented, not reaching posterior margin of cephalothorax.Armature formula: 7.2.5.2.2.2.3.2+A.2.2+A.7+A. First segment ornamented with transverse row of spinules on proximal anterior margin. Fifth segment with spiniform seta. Terminal segment with subapical acrotheck. Aesthetasc on eighth and tenth segments inserted near seta; aesthetasc on eighth segment shorter than seta, on tenth as long as seta.
ANTENNA ( Fig. 7B View Fig. 7 ). 4-segmented; with coxobasis and 3-segmented Endp. Coxobasis with three setae; seta representing Exp not extending beyond Endp-2. Endp-1 with longitudinal row of spinules along distal half of inner (caudal) margin; with single smooth seta on outer margin. Endp-2 about 1.5 times as long as wide, with longitudinal row of minute spinules along inner margin and with five smooth setae along outer margin (three laterally, one subapically, one apically). Endp-3 segment twice as long as wide, with longitudinal row of minute spinules arranged in two rows, armed with seven smooth apical setae (two of them robust and curved).
MANDIBLE ( Fig. 7C View Fig. 7 ). With coxa and basis; gnathobase with strong chitinized teeth; ventralmost teeth robust and obtuse, dorsalmost sharp; pinnate seta dorsally. Mandibular palp 1-segmented, with one short and two very long setae. Two long setae about 8.0 times as long as short seta.
MAXILLULE ( Fig. 7D View Fig. 7 ). With robust praecoxa and 2-segmented palp; proximal segment of palp coxobasis; distal one Endp. Arthrite of praecoxa with six strong spines laterally; proximal one plumose, distal one smooth; apically with thee claw-like spines decreasing in length and one spiniform seta. Coxobasis with three elements; one robust bipinnate seta accompanied by two thin setae apically. Endp with two setae apically and one seta subapically. Exp represented by one seta.
MAXILLA ( Fig. 7E View Fig. 7 ). 5-segmented. Endite of praecoxa prominent, with two pinnate setae. Proximal endite of coxa with one pinnate seta; distal endite elongate, with two setae apically; longest one bipinnate, other one smooth; pinnate seta about 1.5 times as long as smooth one. Basis with claw-like basal endite, with a row of strong spinules along concave margin; distalmost the largest; two setae at base of claw; longest seta as long as claw; other seta 0.5 times as long as previous one. Endp 2-segmented; proximal segment with two robust setae, distal segment with one robust seta apically; as long as claw-like expansion on basis, accompanied by two slender, shorter setae.
MAXILLIPED ( Fig. 7F View Fig. 7 ). 4-segmented; syncoxa and basis with two unipinnate setae each; basis with a row of spinules on distal outer margin. Proximal segment of Endp with one robust, unipinnate seta; distal one armed with three setae; shortest seta smooth.
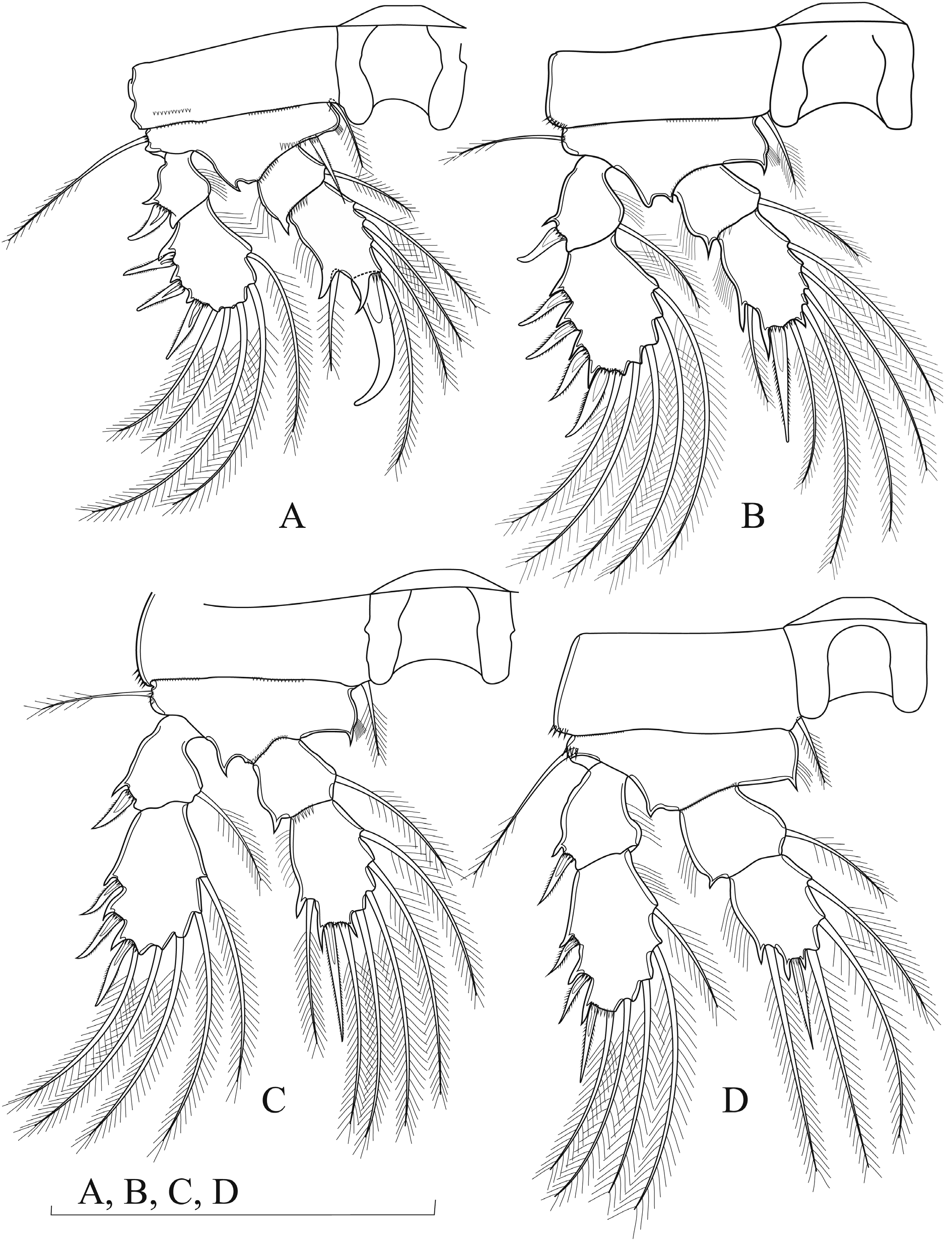
P1–P4. With unornamented intercoxal sclerite, with concave posterior margins; coxa rectangular with a row of spinules on distal outer corner, except P1; basis sub-triangular with slender outer seta with few spinules at implantation ( Fig. 8A–D View Fig. 8 ). Two-segmented Exp and Endp. Exp-2 spine/setal formula: 3.4.3.3/5.5.5.5. Endp-2 spine/setal formula: 1.1.1.1/5.6.6.4. Armature of P1–P4 as follows (Roman numbers = spines; Arabic numbers = setae):
P1 ( Fig. 8A View Fig. 8 ). Exp-1 as long as wide; with short, blunt, curved spine on outer margin. Exp-2 as long as wide. Endp-1 0.7 times as long as wide. Endp-2 1.5 times as long as wide, with one outer seta between two claw-like cuticular extensions; strong, blunt, hook-shaped spine apically.
P2 ( Fig. 8B View Fig. 8 ). Exp-1 slightly longer than wide, with one curved smooth spine on outer corner. Exp-2 about 1.5 times as long as wide. Endp-1 as long as wide. Endp-2 about twice as long as wide; apical spine as long as segment bearing it.
P3 ( Fig. 8C View Fig. 8 ). Similar to P2 but Exp-2 with three spines along outer margin.
P4 ( Fig. 8D View Fig. 8 ). Exp-1 about 1.2 times as long as wide. Exp-2 about 2.2 times as long as wide; spines relatively slim and weak. Endp-1 as long as wide. Endp-2 slightly longer than wide; apical spine as long as segment bearing it.
P5 ( Fig. 6E–F View Fig. 6 ). With baseoendopod fused to Pd5, with one vestigial seta. Distal segment free, wider than long, with unipinnate apical outer seta and short inner spine. Apical seta about 4.0 times as long as inner spine.
P6 ( Fig. 6C View Fig. 6 ). Small, forming simple cuticular plate, inserted laterodorsally on genital double-somite, with three elements; dorsalmost short seta, other two very short, spiniform.
EGG SACS ( Fig. 6A View Fig. 6 ) Extruded from gonopores laterodorsally, with 5–8 eggs in each sac.
Male
Unknown.
Differential diagnosis and remarks
Metacyclops thailandicus sp. nov. differs clearly from the other members of the genus, including the most closely related M. cushae Reid, 1991 . However, both species have some affinities to the genus Hesperocyclops Herbst, 1984 , as well. The following are common characters shared among M. thailandicus sp. nov. from Thailand (South East Asia) and M. cushae from Louisiana (southern part of North America): a) antennules 11-segmented, b) spine formula of swimming legs P1–P4 3.4.3.3, c) P4 Endp-2 with one apical spine, d) intercoxal sclerite of swimming legs without ornamentation, e) apical accessory seta (VI) on caudal ramus shorter than posterolateral seta (III), f) antenna with seta representing Exp not reaching tip of the Endp-2 segment, g) antenna Endp-2 with 5 setae. Three additional species similar to the new species, M. hannensis Defaye, 1992 , M. ryukyuensis Ishida, 1995 , and M. mortoni Pesce, De Laurentiis & Humphreys, 1996 , not included in the differential diagnosis of Reid (1991) between M. cushae and other representatives of the genus, also have one apical spine on P4 Endp-2, but M. hannensis and M. ryukyuensis have the spine the formula 3.4.4.3, while M. mortoni has 12-segmented antennules and spine formula 3.4.4.2.
Metacyclops thailandicus sp. nov. is distinct from M. cushae by the following characters: a) P1 Exp-1 without seta on its inner margin in M. thailandicus sp. nov., b) body without pitting ornamentation in M. thailandicus sp. nov., c) Endp-2 of antenna and the seventh segment of antennules are relatively longer in M. thailandicus sp. nov., d) caudal ramus length/width ratio in M. thailandicus sp. nov. is 2.6 compared to 3.0 in M. cushae , e) and maxilliped with surface ornamentation on syncoxa and basis in M. thailandicus sp. nov., which is absent in M. cushae .
The characteristic spine formula 3.4.3.3 and P4 Endp-2 with one element apically in M. thailandicus sp. nov. are the main differences between it and the rest of the members of the genus, where normally there are two spines apically. Four additional taxa, with one spine apically on P4 Endp-2, previously placed in the Metacyclops trispinosus group ( Karanovic 2004) were recently placed in a new genus, Pescecyclops Karanovic, Eberhard & Murdoch, 2011 , which includes P. kimberleyi ( Karanovic, 2004) , P. laurentiisae ( Karanovic, 2004) , P. pilbaricus ( Karanovic, 2004) and P. pilanus ( Karanovic, 2004) . They differ clearly from M. thailandicus sp. nov. and M. cushae by their spine formula, 3.3.3.3.
Considering the characters of the genus Hesperocyclops described by Herbst (1984) and emended by Galassi & Pesce (1992), it is shown that the characters of M. thailandicus sp. nov. and M. cushae match well with it in: a) spine formula 3.4.3.3, b) apical accessory seta (VI) on caudal ramus shorter than posterolateral one (III), c) antenna with seta representing Exp, d) inner corner of basipodite of P1 with round prominence (well formed in M. cushae and less so in M. thailandicus sp. nov.), e) P5 Exp wider than long, two elements apically inserted far from each other. The significant difference between those two Metacyclops species and those of the genus Hesperocyclops is the modification of P4 Endp in females. In both Metacyclops species, P4 Endp of the female is not modified. The general modification of P4 Endp in Hesperocyclops is an expansion of the proximal segment. In some species, the proximal segment is completely fused with the distal one. On the other hand, normally 2-segmented P4 Endp was found in males of all representatives of the genus Hesperocyclops (Pesce 1985; Dussart & Frutos 1986; Rocha & Carvalho Bjornberg 1987; Galassi & Pesce 1992).
Since the review of Cyclopinae two decades ago ( Pesce 1996) and until now, 21genera and subgenera have been added to the subfamily. So far, representatives of eight genera within the subfamily Cyclopinae have P5 with only one free segment (i.e., Exp), while the baseoendopod is completely fused with the Pd5. They have 2-segmented P1–P4 Exp/Endp and 10/11-segmented antennules. Of those eight genera, three ( Meridiecyclops Fiers, 2001 ; Fierscyclops Karanovic, 2004 ; Pescecyclops Karanovic, Eberhard & Murdoch, 2011 ) were established to accommodate some previous members of the genus Metacyclops Kiefer, 1927 ( Fiers 2001; Karanovic 2004; Karanovic et al. 2011), while some others were synonymised, i.e., M. ( Apocyclops) stocki Pesce, 1985 was recognised as a synonym of Hesperocyclops improvisus Herbst, 1984 ( Galassi & Pesce 1992). This indicates, probably, the polyphyletic nature of the genus Metacyclops and, thus, that a more detailed re-examination of both the P5 and many currently neglected (micro)characters, including sequence data, is urgently needed ( Fiers 2001; Karanovic 2004). Many taxa from the Microcyclops group live in groundwater habitats (caves and interstitial water). Those habitats are characterised by specific ecological conditions, like scarcity of food, constant temperature, absence of light and, in interstitial waters, small spaces. In such habitats, convergent evolution can take place (i.e., through simplifications) ( Sket 1985) and only analyses of gene sequences can resolve real relationships between taxa.
At present, representatives of 25 genera and subgenera of Cyclopinae can be included in the Microcyclops group. It is a morphologically very diverse group, occupying a wide array of habitats from epigean to subterranean. The species of the Microcyclops group are characterized by 10- or 11-segmented antennules, reduced P5, either as a separate segment or being more or less fused with the Pd5, usually as an elongated tubercle-like structure, and with P1–P4 2-segmented. However, there are exceptions: in the genus Hesperocyclops the female P4 Endp is 1-segmented, while in Itocyclops and Rheocyclops Reid & Strayer, 1999 the female Exp is 3-segmented (Reid et al. 1999). The P5 Exp of the following genera has only one single seta: Cryptocyclops G.O. Sars, 1927 ; Neutrocyclops Kiefer, 1936 ; Idiocyclops Herbst, 1987 . The genera with 2 setae/spines on the Exp are Anzcyclops Karanovic, Eberhard & Murdoch, 2011 ; Apalachocyclops Fiers, 2011 ; Apocyclops Lindberg, 1942 ; Cochlacocyclops Kiefer, 1955 ; Dussartcyclops Karanovic, Eberhard & Murdoch, 2011 (with subgenus Barrowcyclops Karanovic, Eberhard & Murdoch, 2011 ); Fierscyclops Karanovic, 2004 ; Fimbricyclops Reid, 1993 ; Goniocyclops Kiefer, 1955 ; Graeteriella Brehm, 1926 (with subgenus Paragraeteriella Rylov, 1948 ); Hesperocyclops Herbst, 1984 ; Itocyclops Reid & Ishida, 2000 ; Menzeliella Lindberg, 1954 ; Meridiecyclops Fiers, 2001 ; Metacyclops Kiefer, 1927 ; Microcyclops Claus, 1893 (inner spine strongly reduced and attached on inner edge or completely lost); Muscocyclops Kiefer, 1937 ; Pescecyclops Karanovic, Eberhard & Murdoch, 2011 ; Pilbaracyclops Karanovic, Eberhard & Murdoch, 2011 ; Psammophilocyclops Fryer, 1956 ; Rheocyclops Reid & Strayer, 1999 ; Teratocyclops Pleşa, 1981 and Speocyclops Kiefer, 1937 .
Key to the genera and subgenera of the Microcyclops group
Common characteristics of the group are the following: a) 10- or 11-segmented antennules, b) P1– P4 with 2-segmented Exp and 1- or 2-segmented Endp (only exceptionally P3–P4 Exp 3-segmented), c) P5 with one free segment (i.e., Exp) or partly fused with the Pd5, with or without baseoendopodal seta.
1. P5 with one free distal segment or fused to Pd5; with one seta apically …………………………2
– P5 with one free distal segment or fused to Pd5; with two setae/spines apically …………………6
2. P5 reduced to tubercle ………………………………………………… Neutrocyclops Kiefer, 1936
‒ P5 with one free distal segment ……………………………………………………………………3
3. Caudal ramus with three terminal setae ………………………………… Idiocyclops Herbst, 1987
– Caudal ramus with four terminal setae ……………………………………………………………4
4. P4 Endp-2 with two apical spines, outer one much shorter and weaker than the other; coxal seta on P1–P4 thin and small; basis of P2–P4 with row of tiny spines on inner expansion …… ……………………………………………………………………… Cryptocyclops G.O. Sars, 1927
– P4 Endp-2 with two well developed apical spines (exceptionally one spine); coxal seta on P1– P4 clearly overreaching basis; basis of P2‒P4 with fine hairs on inner expansion ………… ………………………………………………………………………… Microcyclops Claus, 1893 (5)
5. P4 Endp-2 with two apical spines ……………………… Microcyclops ( Microcyclops) Claus, 1893
– P4 Endp-2 with one apical spine …………… Microcyclops ( Mediocyclops) Dussart & Sarnita, 1986
6. Pd3–Pd5 with rows of long spinules laterally ………………………… Fimbricyclops Reid, 1993
– Pd3–Pd5 with no rows of long spinules laterally ………………………………………………7
7. Pd5 without lateral seta (vestige of the basis of P5) ………………… Graeteriella Brehm, 1926 (8)
– Pd5 with lateral seta ………………………………………………………………………………9
8. Caudal ramus with five setae; P4 Exp with 2-segmented ramus …………………………………… …………………………………………………………… Graeteriella ( Graeteriella) Brehm, 1926
– Caudal ramus with six setae; P4 Exp with 3-segmented ramus …………………………………… ………………………………………………………… Graeteriella ( Paragraeteriella) Rylov, 1948
9. Proximal half of genital double-somite with two sclerotized external parallel crests ventrally; inner margin of basis and P4 Endp-1 expanded ………………………………… Meridiecyclops Fiers, 2001
‒ Genital double-somite without modified structures ventrally; inner margin of basis with or without expansion; P4 Endp-1 without expansion …………………………………………………………10
10. Female P3–P4 Exp 2-segmented …………………………………………………………………12
– Female P3–P4 Exp 3-segmented; if P4 Exp 2-segmented, anal operculum not extending beyond articulation of caudal ramus ……………………………………………………………………11
11. Anal operculum well developed, irregularly triangular shaped; reaching beyond articulation of caudal ramus; A2 Endp-2 with eight setae ……………………………… Itocyclops Reid & Ishida, 2000
– Anal operculum short, not reaching beyond articulation of caudal ramus; A2 Endp-2 with six or seven setae ……………………………………………………… Rheocyclops Reid & Strayer, 1999
12. Male P3–P4 Exp 3-segmented; number of segments of P3–P4 Exp sexual dimorphic character ………………………………………………………………………… Apalachocyclops Fiers, 2011
– Male P3–P4 Exp 2-segmented; number of segments of P4 Endp not sexual dimorphic character …13
13. P5 with considerable distance between two elements apically …………………………………14
– P5 with spine and seta inserted close to each other; if P5 with considerable distance between two elements apically, female P4 Endp similar in shape to other swimming legs ………………………15
14. Female P4 Endp 2-segmented; with one spine and one seta apically …… Apocyclops Lindberg, 1942
‒ Female P4 Endp 1- or 2-segmented, conspicuously modified, very different from that of male; with one spine apically …………………………………………………… Hesperocyclops Herbst, 1984
15. P5 with free distinct segment; with one robust, blade-like spine and one seta; spine on distal segment of P5 as long as seta; anterior part of caudal ramus with a row of spinules dorsoventrally ………………………………………………………… Menzeliella Lindberg, 1954
– P5 with or without free distinct segment; apical elements on segment with two setae or one seta and one spine; caudal ramus without row of spinules dorsoventrally ……………………………16
16. Anal operculum well developed; free margin serrated …………………………………………17 – Anal opeculum well developed; free margin smooth ……………………………………………18
17. Spine formula 3.4.4.3 …………………………………………………… Speocyclops Kiefer, 1937 – Spine formula 2.3.3.2 ………………………………………………… Muscocyclops Kiefer, 1937
18. P2 Exp-2 with four spines; if three spines, P3 Exp-2 with three spines and P4 Endp-2 with two elements apically …………………………………………………………………………………19
– P2 Endp-2 with three or two spines ………………………………………………………………21
19. Genital double-somite barrel-shaped; P4 Endp-2 with one slender, setiform spine apically, twice as long as segment bearing it; outer apical seta (IV) on caudal ramus with geniculation (to prevent breaking) ………………………………………………………………… Teratocyclops Pleşa, 1981
– Characters not as above ……………………………………………………………………………20
20. P5 with robust spine and short seta, spine about 0.5 times as long as seta; caudal ramus with large anterolateral accessory seta (I) …………………………… Fierscyclops Karanovic, 2004
– P5 with spine and long seta, spine less than 0.5 times as long as seta; caudal ramus without anterolateral accessory seta (I) ………………………………………… Metacyclops Kiefer, 1927
21. Caudal ramus with large anterolateral accessory seta (I) …………………………………………… …………………………………………… Pilbaracyclops Karanovic, Eberhard & Murdoch, 2011
– Caudal ramus without anterolateral accessory seta (I) …………………………………………22
22. P1 Exp-2 with three spines; if two spines caudal ramus more than twice as long as wide …………23 – P1 Exp-2 with two spines …………………………………………………………………………25
23. Caudal ramus more than three times as long as wide; if less, distal segment of P5 free; with one spine and one seta apically ………………… Pescecyclops Karanovic, Eberhard & Murdoch, 2011
– Caudal ramus less than 2.6 times as long as wide; if more, distal segment of P5 fused to Pd5; with two setae apically …………………………………………………………………………24
24. Seminal receptacle well developed, three foiled, posterior part circular; spine formula 3.3.3.2 …… ……………………………………………………………………… Cochlacocyclops Kiefer, 1955
– Seminal receptacle less developed posteriorly; spine formula 3.3.3.2, 3.3.3.3 or 2.2.2.2 ……… …………………………………………………………………………… Goniocyclops Kiefer, 1955
25. P2 Endp-2 with three spines …………………… Anzcyclops Karanovic, Eberhard & Murdoch, 2011 – P2 Endp-2 with two spines …………… Dussartcyclops Karanovic, Eberhard & Murdoch, 2011 (26)
26. P5 distal segment free …… Dussartcyclops ( Dussartcyclops) Karanovic, Eberhard & Murdoch, 2011
– P5 distal segment fused to Pd5 ……………………………………………………………………… ………………………… Dussartcyclops ( Barrowcyclops) Karanovic, Eberhard & Murdoch, 2011
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cyclopinae |
|
Genus |