
Sesarmidae Dana, 1852
|
publication ID |
https://doi.org/10.26107/RBZ-2020-0097 |
|
publication LSID |
lsid:zoobank.org:pub:815E4670-B063-4FD8-B31E-3AD89B3A7942 |
|
DOI |
https://doi.org/10.5281/zenodo.6525275 |
|
persistent identifier |
https://treatment.plazi.org/id/49152B56-FFE2-BA27-FC64-FD4DFBCEFBA4 |
|
treatment provided by |
Diego |
|
scientific name |
Sesarmidae Dana, 1852 |
| status |
|
Family Sesarmidae Dana, 1852 View in CoL
Historical and nomenclatural issues associated with
Chiromantes and allied genera. Chiromantes Gistel, 1848 , has had a long and often confused history that is not restricted to the species level. Until 40 years ago, members of Chiromantes were still classified in Holometopus H. Milne Edwards, 1853 , but Holthuis (1977: 170) argued that their respective type species had been incorrectly assigned. He showed that both Chiromantes Gistel, 1848 , and Holometopus H. Milne Edwards, 1853 , share the same type species, Grapsus ( Pachysoma) haematocheir De Haan, 1833 , and as such, were objective synonyms, with Chiromantes Gistel, 1848 , having priority. Holthuis (1977: 170) also noted that all the species that had been assigned to Chiromantes by workers up to that point should instead be referred to Perisesarma De Man, 1895 ( type species Sesarma dusumieri H. Milne- Edwards, 1853, subsequent designation of Campbell, 1967).
Serène & Soh (1970) completely reorganised the generic and subgeneric classification of the Indo-West Pacific Sesarmidae (when still considered a subfamily, Sesarminae ) and established many new genera and subgenera. However, they did not change the concept of what had previously been the subgenus Chiromantes (as Holometopus ). Chiromantes has continued to include all those species with an entire lateral carapace margin (no trace of an epibranchial tooth), the dorsal margin of the cheliped palm with no (or only one oblique) pectinated ridge, and orbits that include the antennal peduncle ( Serène & Soh, 1970: 388–392). Chiromantes thus remained one of the larger and better-known Indo- West Pacific sesarmid genera. In his 1968 checklist, Serène recognised 13 species (under subgenus Holometopus ): C. boulengeri Calman, 1920 , C. dehaani (H. Milne Edwards, 1853) , C. elongatus (A. Milne-Edwards, 1869) , C. eulimene (De Man, in Weber, 1897), C. eydouxi (H. Milne Edwards, 1853) , C. granosimanum ( Miers, 1880) , C. haematocheir ( De Haan, 1833) , C. obesus ( Dana, 1851) , C. obtusifrons ( Dana, 1851) , C. ortmanni ( Crosnier, 1965) , C. stormi ( De Man, 1895) , C. tangi ( Rathbun, 1931) , and C. villosus (A. Milne-Edwards, 1869) . Subsequently, Soh (1978) described a new species, C. serenei , from Hong Kong.
An appraisal of Chiromantes by Ng & Liu (1999) noted that two major groups could be discerned in Chiromantes . One group had a single ridge of tubercles on the dorsal margin of the palm and differentiated granules on the dorsal margin of the dactylus. Included species were C. elongatus and C. villosus (both with a single longitudinal pectinated ridge), C. eydouxi (with one longitudinal tuberculate ridge), and C. eulimene and C. ortmanni (both with an oblique to transverse single pectinated ridge). Members of the second group, also including C. tangi and C. stormi , have no ridge on the dorsal margin of the palm and the dorsal margin of the dactylus has no, or only weakly differentiated, granules. These two species were discussed in detail by Ng & Liu (1999), with C. tangi provisionally retained in Chiromantes , while C. stormi was referred to a new genus, Stelgistra Ng & Liu, 1999 . Significantly, Ng & Liu (1999: 230) commented (but did not elaborate) that “ C. haematochir [sic] is distinct in several features (especially in the structure of the anterior male thoracic sternum) and should be generically separated from the other species in the group”. Since C. haematocheir is the type species of Chiromantes , this would imply that all the other species in the genus would need to be moved to other genera.
Ng & Liu (1999) referred Sesarma villosum to Clistocoeloma A. Milne-Edwards, 1873 and suggested that Sesarma obesum could be a species of Metasesarma H. Milne Edwards, 1853 . In a molecular study, Schubart et al. (2006: 197) supported the placement of Sesarma villosum within Clistocoeloma . Ng & Schubart (2003) showed that Sesarma obesum Dana, 1851 , is a senior subjective synonym of Metasesarma rousseauxi H. Milne Edwards, 1853 , and designated a neotype for Dana’s species. They also demonstrated that Sesarma eydouxi H. Milne Edwards, 1853 (type locality supposedly Vietnam), is not a species of Chiromantes , but a junior synonym of the American Sesarma rectum Randall, 1840 . Schubart & Ng (2002) transferred Chiromantes tangi ( Rathbun, 1931) , to Neosarmatium Serène & Soh, 1970 , noting that it is close to N. laeve (A. Milne-Edwards, 1869) . A recent reappraisal of C. tangi by Ng et al. (2019), however, showed that it is not closely related to Neosarmatium , and was transferred to its own genus, Sinosesarma .
Some results of the present study have been foreshadowed by Ng et al. (2008a: 223–225), notably in regard to the taxonomic complexities of Bresedium , Chiromantes , Pseudosesarma , and Sesarmops . Ng et al. (2008a) listed eight species in Chiromantes : C. boulengeri Calman, 1920 , C. dehaani (H. Milne Edwards, 1853) , C. eulimene (De Man, in Weber, 1897), C. haematocheir ( De Haan, 1833) , C. neglectum ( De Man, 1887) , C. obtusifrons ( Dana, 1851) , and C. ortmanni ( Crosnier, 1965) ; placing Sesarma granosimana Miers, 1880 , provisionally in Pseudosesarma Serène & Soh, 1970 , but without explanation. However, they added on the observations of Ng et al. (2001: 41) that Sesarma neglecta De Man, 1887 , was a valid species allied to C. dehaani . Ng et al. (2008a) also noted that Sesarma serenei Soh, 1978 , should be considered a junior subjective synonym of C. haematocheir (see treatment in Naruse & Ng, 2008). Ng et al. (2008a) commented that ongoing revisions of Chiromantes and related taxa, like Sesarmops and Pseudosesarma , would substantially change the generic classification of the species in these genera. In fact, the only species they anticipated to remain in Chiromantes sensu stricto (s. str.) was Sesarma haematocheir De Haan, 1833 . Naruse & Ng (2008) later described C. ryukyuanus as a sister species of C. haematocheir from the Ryukyu Islands in Japan. In their revision of a number of Indo-Pacific taxa with a longitudinal pectinated crest, Schubart et al. (2009) referred Chiromantes elongatus (A. Milne-Edwards, 1869) to Selatium Serène & Soh, 1970 . Comparative morphological and genetic studies also revealed that some representatives of Pseudosesarma and Sesarmops have close affinities to Chiromantes s. lat. and would need to be revised simultaneously.
With regard to Pseudosesarma, Serène & Soh (1970) established the genus for species with the following characteristics: a) medium-sized species with an epibranchial tooth and the antennal peduncle being inside the orbit, but without pectinated crests on male chelar palm; b) carapace slightly convex and shorter than wide, with lateral border of carapace weakly or not diverging posteriorly; c) anterior frontal margin with feeble median concavity, postfrontal lobes not remarkably salient. They made Sesarma edwardsii De Man, 1887 , the type species and included seven species: Sesarma bocourti A. Milne-Edwards, 1869 , Sesarma crassimanum De Man, 1887 , Sesarma edwardsii De Man, 1887 (spelled incorrectly as “ edwarsi ”), Sesarma johorensis Tweedie, 1940 , Sesarma laevimanum Zehntner, 1894 , Sesarma modestum De Man, 1902 , and Sesarma moeschii De Man, 1892 . Soh (1978) added a new species, Pseudosesarma patshuni , from Hong Kong.
Ng et al. (2008a: 222, 225) listed nine species in Pseudosesarma , thereby including P. granosimanum ( Miers, 1880) , a species previously classified in Chiromantes due to the lack of anterolateral carapace teeth, without detailed elaboration (see above). They noted that Pseudosesarma crassimanum , P. johorense , P. moeschii , and P. patshuni may need to be referred to other genera in the future (see also Ng & Schubart, 2017). The present revision of Chiromantes thus requires the simultaneous reappraisal of Pseudosesarma .
Serène & Soh (1970) clearly had reservations about their generic assignments of several species, recognising some of the unresolved generic problems. In their remarks on Pseudosesarma , they wrote: “The genus has several characters close to those of Sesarmops and the separation of the two genera needs to be improved. As it is, Pseudosesarma is still heterogeneous. Perhaps a new genus would have to be established giving priority to the shape of the male pleopod and grouping bocourti with the species of Sesarmops which, like intermedium have the same type of male pleopod. The species moeschi and johorensis with their smooth shining carapace and other characters are also a few aberrant in[to] Pseudosesarma ” ( Serène & Soh, 1970: 400). As these issues have never been resolved, it is necessary to include Sesarmops Serène & Soh, 1970 , as part of the present revision.
Sesarmops was established for six species: Sesarma atrorubens Hess, 1865 , Sesarma impressus H. Milne Edwards, 1837 ( type species by original designation), Sesarma intermedius De Haan, 1835 , Sesarma mindanaoensis Rathbun, 1914 , Sesarma sinensis H. Milne Edwards, 1853 , and Sesarma weberi De Man, 1892 . Ng et al. (2008a) continued to include these species, but commented that Sesarmops intermedius , S. sinensis , and S. weberi need to be transferred to other genera. This was already noted by Serène & Soh (1970: 401): “If priority is given to the shape of the male pleopod, intermedium , sinensis can hardly be considered as congeneric with impressum . However, the type of the male pleopod of weberi is identical with that of intermedium and the fact that De Man (1902) considered intermedium as a synonym of impressum is at least an indication to support the present position. S. sinensis by its carapace clearly shorter than breadth between the external orbital angles and with lateral border nearly parallel seem to be aberrant in the genus. At least by the ornamentation of the dactylus of male cheliped weberi is somewhat aberrant into Sesarmops ”.
The present study demonstrates that Pseudosesarma patshuni , Sesarmops intermedius , and S. sinensis are morphologically and phylogenetically close to Chiromantes dehaani and C. neglectus , while Pseudosesarma moeschii and P. johorense share a unique set of morphological characters. Sesarma edwardsi var. laevimana Zehntner, 1894 (often placed in Pseudosesarma ) is here recognised as the senior synonym of Bresedium sediliense ( Tweedie, 1940) , and remains in Bresedium for the moment (see Remarks for that genus). Pseudosesarma s. str. is here restricted to P. edwardsii , P. crassimanum , and allies, now also including Chiromantes boulengeri (see Ng & Schubart, 2017). Pseudosesarma bocourti and P. granosimanum are referred to two new genera.
With the present reorganisation and recent reappraisal of the type species by Ng et al. (2020), Sesarmops now contains S. angustifrons (A. Milne-Edwards, 1869) , S. atrorubens ( Hess, 1865) , S. imperator Ng, Li & Shih, 2020 , S. impressus (H. Milne Edwards, 1837) , S. indicus Ng, Li & Shih, 2020 , S. mindanaoensis ( Rathbun, 1914) , S. similis Hess, 1865 , and S. weberi ( De Man, 1892) (see also Paulay & Starmer, 2011; Naruse & Ng, 2020). The genus is not monophyletic and will be revised at a later date.
The new taxonomic structure provided here, including the nine new genera proposed, is based on newly recognised morphological characters, supported by a phylogenetic analysis, and clearly shows that the presence or absence of an epibranchial tooth is not useful in systematics (see General Discussion).
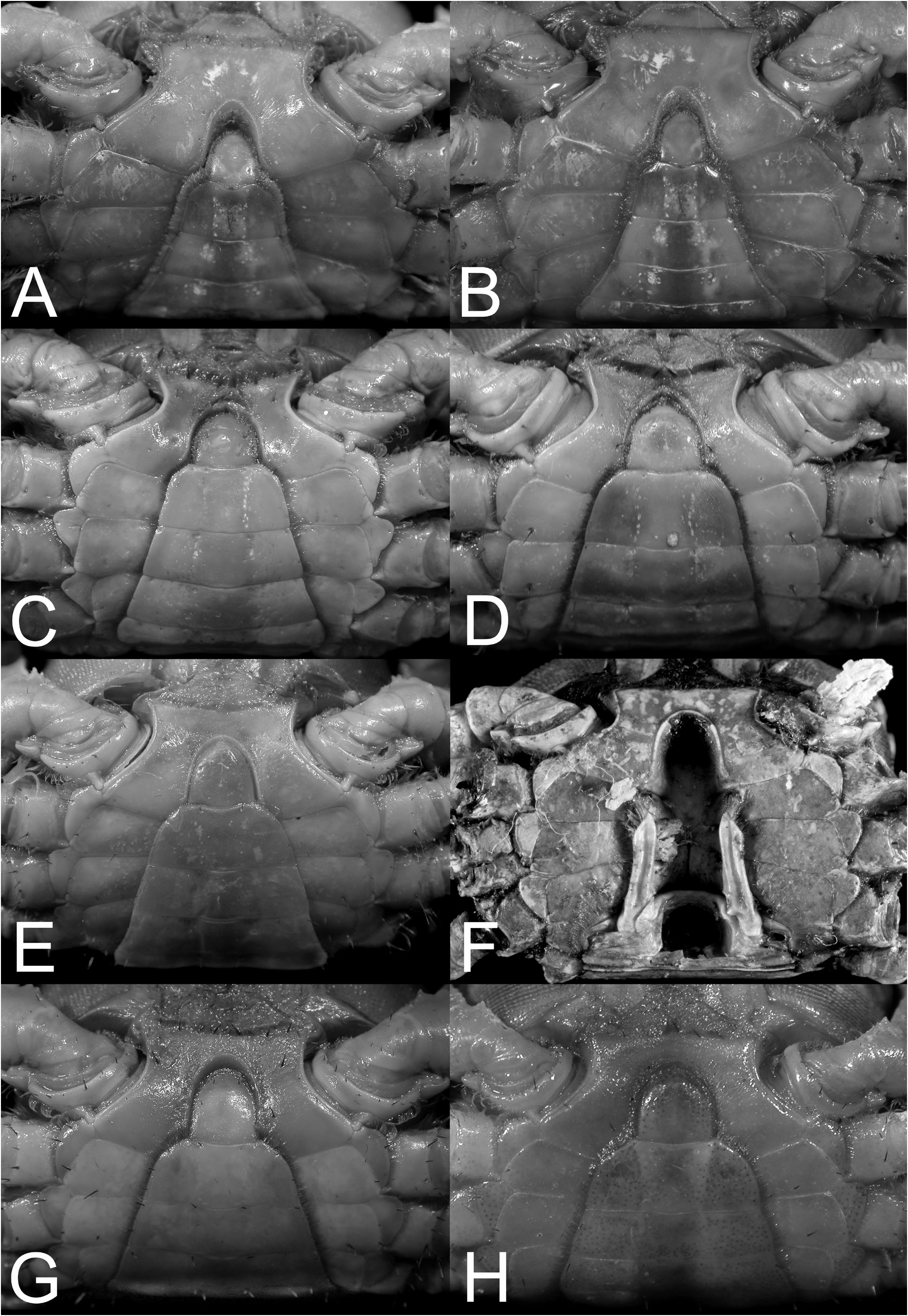
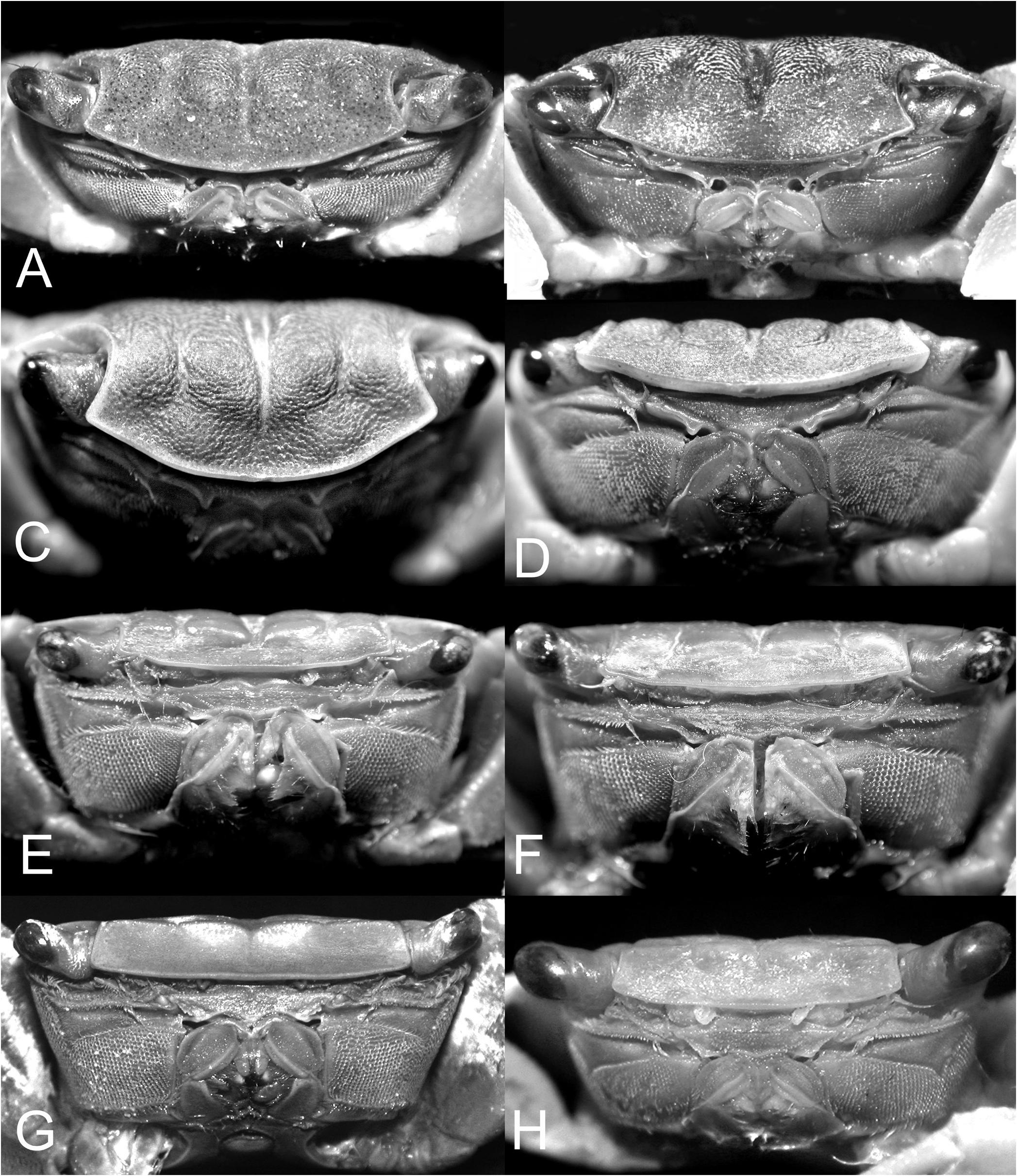
Outline of the new generic structure. The type species of Chiromantes , Grapsus ( Pachysoma) haematocheir De Haan, 1833 , is markedly different from the other species currently placed in the genus, with the exception of C. ryukyuanus and the already synonymised C. serenei , because its adult male thoracic sternites 2 to 4 are relatively broader and the sternopleonal cavity reaches only to the median point of sternite 4 ( Fig. 9A, B View Fig ). All other species assigned to Chiromantes have thoracic sternites 2 to 4 proportionately narrower and the sternopleonal cavity longer, reaching to just before the anterior margin of sternite 2 ( Fig. 9C–H View Fig ).
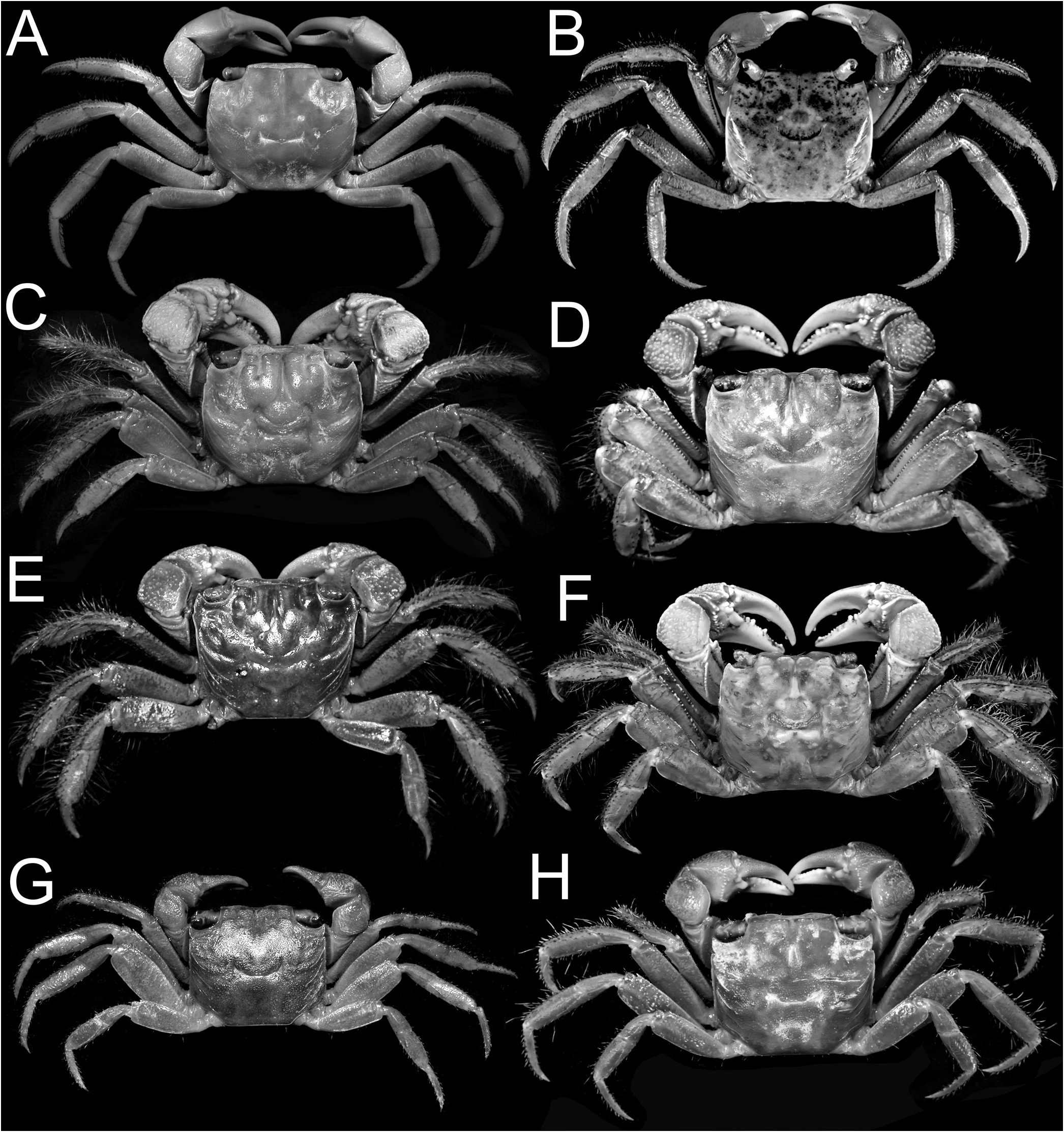
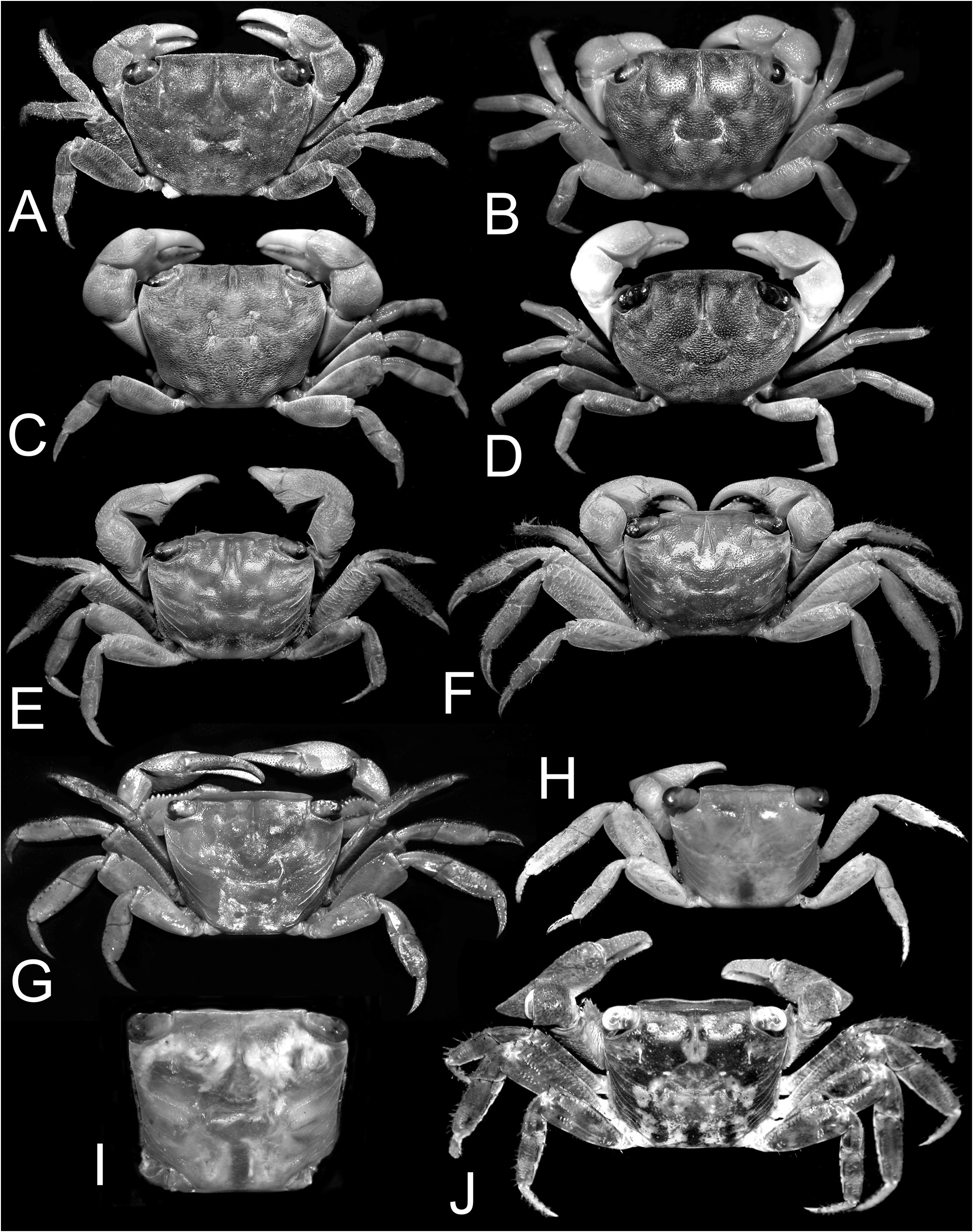
With Chiromantes haematocheir and C. ryukyuanus separated, the remaining species fall into several discrete groups that will be given genus rank: Chiromantes eulimene (De Man, in Weber, 1897) and C. ortmanni ( Crosnier, 1965) are in one group, while C. dehaani (H. Milne Edwards, 1853) , C. neglectum ( De Man, 1887) , and C. magnus Komai & Ng, 2013 , are in another. Chiromantes obtusifrons ( Dana, 1851) and four species described as new in the revision of this group of species by Davie & Ng (2013) belong to a third group. Chiromantes angolensis ( Brito Capello, 1864) and C. buettikoferi ( De Man, 1883) belong to their own respective groups. Chiromantes boulengeri Calman, 1920 , belongs in Pseudosesarma , while C. granosimanum ( Miers, 1880) possesses sufficient unique morphological characters to justify its placement in a monotypic genus. Similarly, Pseudosesarma bocourti (A. Milne-Edwards, 1869) is also referred to a new genus, and its former synonym Sesarma cheirogona Targioni Tozzetti, 1877 , is here regarded as a valid species.
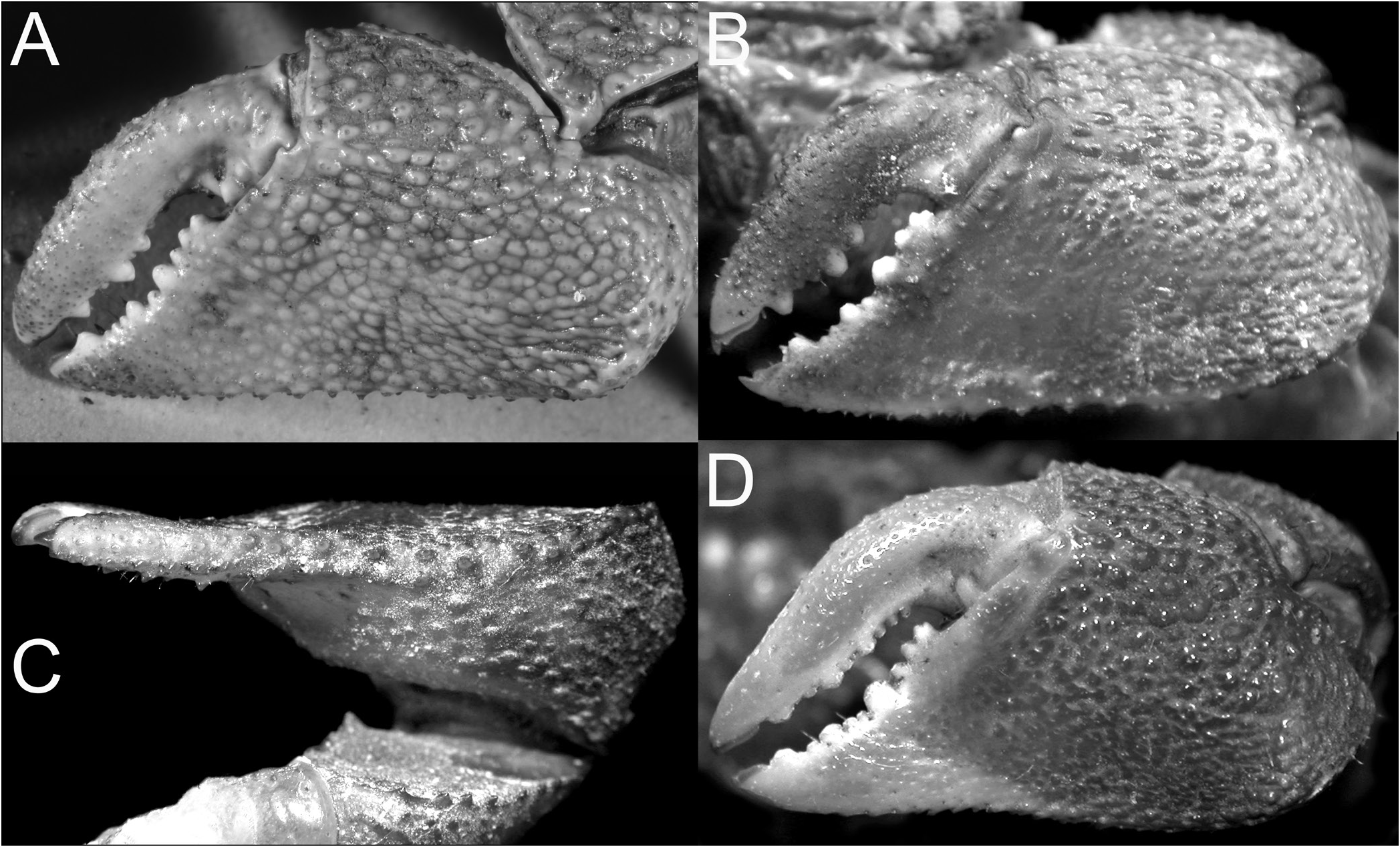
Chiromantes dehaani s. str., C. neglectus , and C. magnus are characterised by their chelipedal palm lacking a longitudinal ridge on its dorsal margin and the dactylus not possessing any distinctly shaped or regularly arranged granules or tubercles along its dorsal margin. On the other hand, the inner surface of the male palm has a prominent transverse ridge of rounded granules that are positioned on the most swollen part of the palm; and the outer surface of the palm has a clearly discernible short and smooth median longitudinal ridge (although sometimes low or partially obscured by a granular surface). In addition, these species have a septum separating the basal antennal and antennular articles (absent in C. haematocheir and C. ryukyuanus ). The structure of the cristae on the postfrontal margin also differs in the two groups. In C. haematocheir and C. ryukyuanus , the epigastric and postfrontal cristae are sharp and almost confluent, with shallow grooves separating them ( Figs. 1A, B View Fig , 3A View Fig ). In C. dehaani s. str., C. neglectus and C. magnus , the epigastric and postfrontal cristae are rounded and separated by deep and broad grooves (e.g., Figs. 1C, D View Fig , 3B View Fig ). In addition, the vulvae are slightly different, with that of C. haematocheir and C. ryukyuanus just touching the margin of sternite 5 ( Fig. 43A View Fig ), while in C. dehaani s. str., C. neglectus , and C. magnus , the vulva is pressed against sternite 5, causing a slight indentation ( Fig. 43B View Fig ). As such, these three species are here referred to their own new genus, Orisarma . This new genus will also accommodate two species that had previously been placed in Sesarmops ( S. sinense and S. intermedius ) and one from Pseudosesarma ( P. patshuni ). The characters noted above for C. dehaani s. str., C. neglectus , and C. magnus also apply to them, including the form of the vulva ( Fig. 43C, D View Fig ), but the epigastric and postorbital cristae are less prominently rounded and the grooves more narrow in Sesarmops sinensis and S. intermedius ( Fig. 2I, J View Fig ).
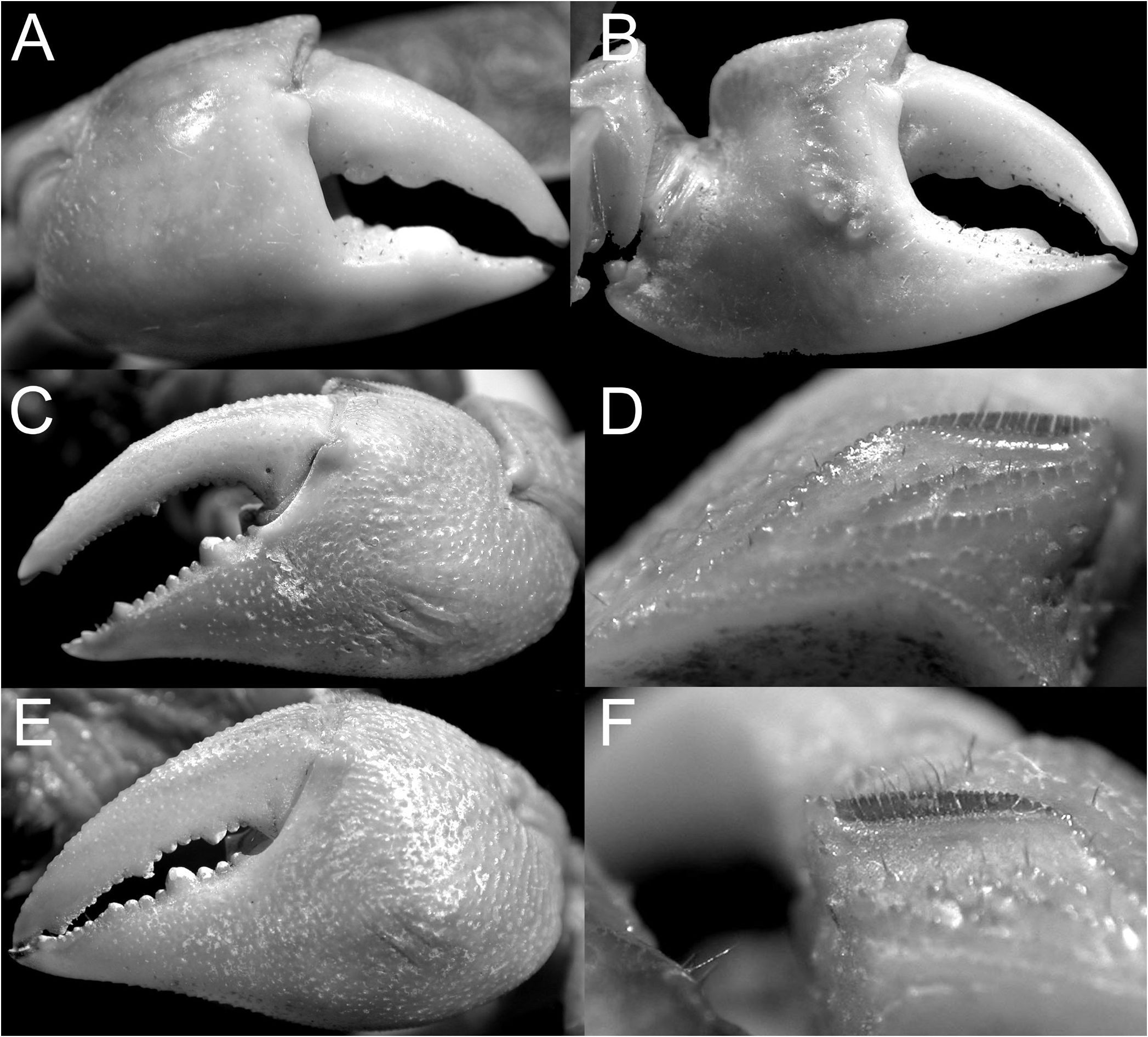
The East African C. eulimene and C. ortmanni are unique in that the dorsal margin of the male palm has a distinct oblique pectinated ridge and the dorsal margin of the chelar dactylus has differentiated granules, presumably for stridulation. These characters are consistent and are here considered to be generically significant. To this effect, we propose to establish a new genus, Cristarma , for these two species. With regard to these two characters, Cristarma is similar to the two known species of Selatium Serène & Soh, 1970 , viz. S. brockii ( De Man, 1887) and S. elongatum (A. Milne-Edwards, 1869) (cf. Schubart et al., 2009). Cristarma can easily be distinguished from species of Selatium in that the pectinated ridge on the male palm is markedly oblique to almost transverse in orientation ( Fig. 15D, F View Fig ) (versus longitudinal), the outer lower surface of the palm has two short, smooth, submedian oblique ridges ( Fig. 15C, E View Fig ) (versus absent), the lateral margins of the ambulatory dactyli are not lined by dense short black setae ( Fig. 13E, F View Fig ), and the surfaces between the first to third ambulatory coxae have dense tufts of long setae ( Fig. 18C, D View Fig ) (versus very short, scattered, or no setae). In addition, the vulva is very low, with the two sternal vulvar covers bracketing the non-projecting opening ( Fig. 43G, H View Fig ). Our molecular phylogenetic tree also shows that there is no close relationship between Selatium and Cristarma or those genera with two transverse pectinated crests, e.g., Parasesarma and Perisesarma ( Fig. 59 View Fig ).
The other two species endemic to Africa, but from the Atlantic coast, C. angolensis and C. buettikoferi , are referred to the new genera, Trapezarma and Platychirarma , respectively. Like Chiromantes s. str., their epigastric and postfrontal crests are sharp and almost continuous ( Figs. 13G, H, J View Fig , 14G, H View Fig ), a character shared with no other group of species. Unlike the West African C. eulimene and C. ortmanni that have pectinated crests on their chelae and granules on the dactylar fingers, the chelae of C. angolensis and C. buettikoferi are smooth and not specially ornamented. What is unusual about their adult male chelae is that the outer surface is gently convex to flat (including the pollex) ( Figs. 16 View Fig , 17 View Fig ), being most extreme in C. buettikoferi . The prominently flattened outer surface of the chela of C. buettikoferi ( Fig. 17 View Fig ) is a character shared only with Pseudosesarma bocourti and C. cheirogonum from Southeast Asia ( Fig. 28 View Fig ) (both referred to a new genus, see later). The carapaces of C. angolensis and C. buettikoferi are quite distinct with a more trapezoidal appearance, with a wider frontal margin and strongly converging lateral margins towards the posterior carapace margin ( Fig. 13G, H, J View Fig ). While the two species are genetically defined as sister species (despite long individual branches, Fig. 59 View Fig ), the suite of cheliped, male pleonal and G1 characters argue against including them in the same genus (see discussion under Trapezarma ). This gains further support from the very differently shaped vulvae, those of Trapezarma widely spaced and the projecting opening directed obliquely posteriorly ( Fig. 43I View Fig ), while the ones of Platychirarma are closer together and the slightly projecting opening is directed obliquely anteriorly ( Fig. 43J View Fig ).
Ng & Liu (1999: 230) had noted that the generic placement of Chiromantes obtusifrons should be reappraised. The trapezoidal carapace of this species is diagnostic, although it shares other characters with Orisarma , new genus. However, C. obtusifrons has a peculiar frontal margin that is very broad and bends downwards so strongly that the antennae and antennules are effectively covered ( Fig. 14A–D View Fig ), the outer surface of the palm has no longitudinal ridge ( Fig. 15A View Fig ), and the surfaces between the first to third ambulatory coxae have distinct dense tufts of long setae ( Fig. 18A, B View Fig ) (a character otherwise only present in C. eulimene and C. ortmanni ). In addition, the vulvae have a totally different structure, with the sternal vulvar covers being plate-like and the opening not projecting ( Fig. 43E, F View Fig ). Davie & Ng (2012) re-examined Chiromantes obtusifrons and recognised four new closely allied species from the Indian Ocean and Western Pacific, and also commented on their generic status. All five species are here referred to a new genus, Danarma .
Ng & Liu (1999) had already described Sesarma ( Sesarma) stormi De Man, 1895 , in detail and explained at length why it should be placed in its own genus Stelgistra Ng & Liu, 1999 . There is no need to elaborate here, as all the characters discussed then are still valid in defining Stelgistra . Stelgistra stormi has since been found in Guam ( Paulay et al., 2003: 508).
The revised classification of the above-discussed species is summarised in Table 2 View Table 2 .







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |