
Cylindromacrostomum formosae, Schockaert, Ernest R., 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3872.5.8 |
|
publication LSID |
lsid:zoobank.org:pub:073E1555-ACF1-40AE-9CE4-342D256B8AE8 |
|
DOI |
https://doi.org/10.5281/zenodo.6136592 |
|
persistent identifier |
https://treatment.plazi.org/id/4404B347-FFD5-F442-FF32-F39CFDC630E7 |
|
treatment provided by |
Plazi |
|
scientific name |
Cylindromacrostomum formosae |
| status |
sp. nov. |
Cylindromacrostomum formosae n.sp.
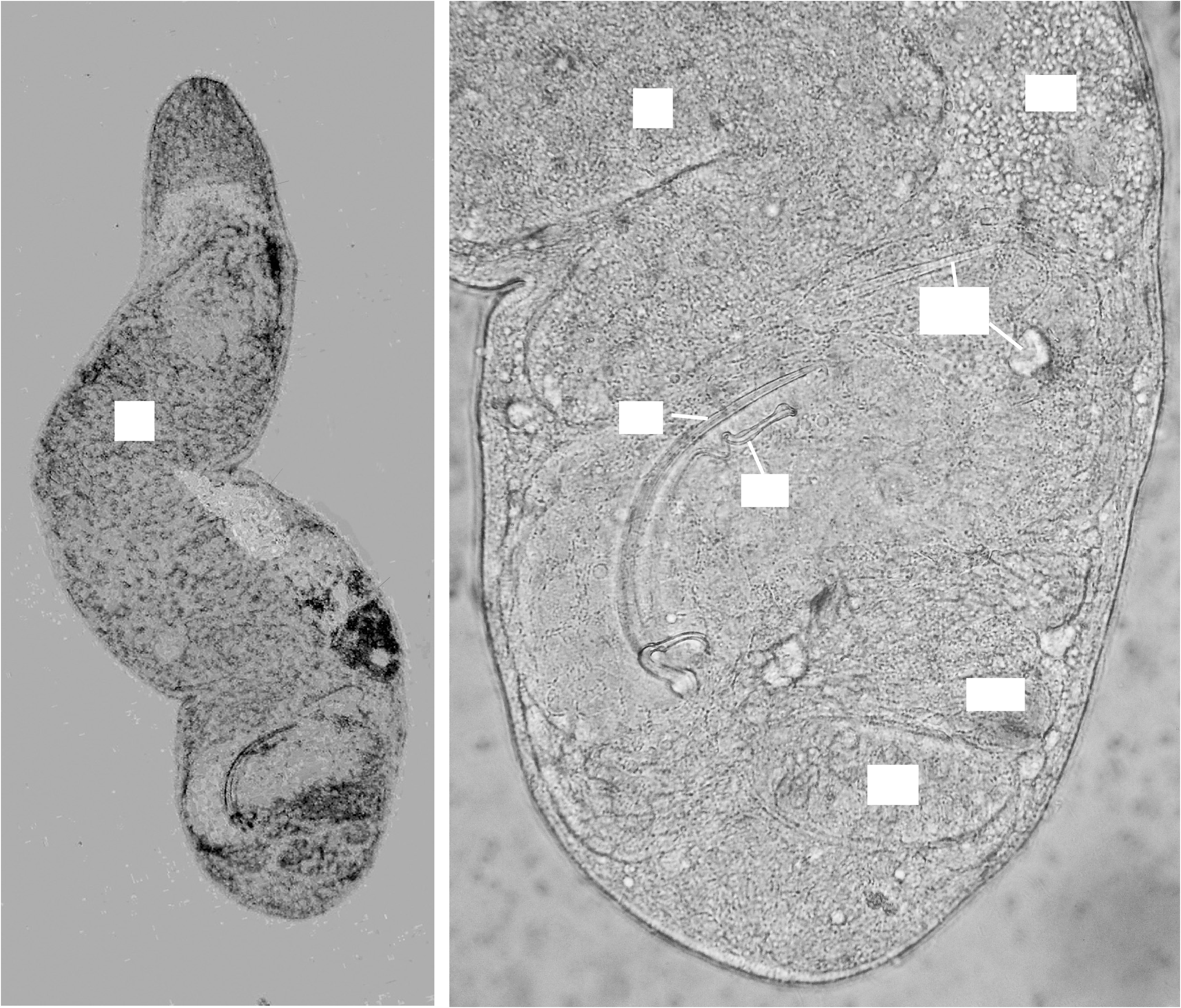
Fig. 5 View FIGURE 5
Localities in the Algarve. Faro ( Portugal), Ria Formosa. Lower mid littoral of a sand ridge with loose and coarse sand with much large shell debris and rather little silt in two similar localities ( type locality 37° 00.392 N, 7° 56.619 W and 36° 59.881 N, 7° 55.673 W on 7 October 2013)
Material. Four whole mounts, two from each location, one of which is designated the holotype ( SMNH. Type- 7840͵ HU nrs PT 561–563); (one whole mount is of a juvenile with only the most distal part of the penis stylet developed), one micrograph of a free-swimming individual at low magnification.
Diagnosis. species of Cylindromacrostomum of about 2 mm, with a penis stylet of 119–129 µm length, the part extending from lateral lamella to the end being 43–46 µm long, a short terminal part that ends bluntly, only slightly narrower than the stylet diameter. The accessory stylet is 71–80 µm. The female hard parts consist of a sperm tube 70–76 µm long, a reniform middle piece 20x26 µm with teeth-like lateral knobs, and a mouthpiece 28 µm long.
Derivation of the name. formosae Latin for ‘of the (Ria) Formosa’.
One specimen is almost 2 mm long in the whole mount. The micrograph of the living animal is unfortunately out of focus but a number of structures are clearly visible including the position of the eyes and the paired ovaries. The big cup-like eyes are much closer to each other than to the lateral sides. In the three adult specimens, the penis stylet is 119–129 µm long (mean = 123 µm), with a constant diameter of around 3–4 µm. As in the two described species of Cylindromacrostomum , C. notandum (Ax, 1951) and C. mediterraneum ( Ax, 1955) , the wall at the proximal opening is thickened, and from there the penis stylet turns around the proximal end of the accessory stylet over about 180°, bends over 45–60° to run, moderately curling, more or less parallel to the accessory stylet. At the last quarter, there is the typical ‘lamella’, which is about 25 µm long and 15 µm wide. Contrary to what we see in most other species, this lamella is completely transparent. There is no turn of 180° before this lamella, as is often the case in species of Dolichomacrostominae (compare, e.g., with Fig. 3 View FIGURE 3 C). At the very end, the penis stylet bends over 90°, narrows very little and ends after 6–7 µm at the terminal opening with a diameter of 2–3 µm. This end part is also surrounded by a thin ‘lamella-like’ sheath. The accessory stylet is 71–80 µm (mean = 77 µm) long (62–69 µm when measured along a straight line), with a proximal opening of 12 µm diameter, and tapers to a sharp end with a lateral opening; in one of the whole mounts, the prostate vesicle is clearly visible and is 120 µm long and 50 µm wide; the canal leading to the penis stylet is 25 µm long.
The hard parts in the female system consist of a single ‘sperm tube’ 70–76 µm long, a kidney-shaped middle piece of 20x26 µm and a mouthpiece 28 µm long. The middle piece is a very thin plate with teeth-like knobs on the border opposite to where sperm tube and mouthpiece meet. There is no interruption between the sperm tube and mouthpiece, which is surrounded by a thin mantle. Where the mouthpiece enters the insemination duct, there are the usual two umbrella-like hard parts. In the holotype the sperm tube starts in a sperm filled vesicle.
This species is a member of the taxon Cylindromacrostomum Rieger, 1971 , witnessed by the presence of eyes and the paired ovaries. Rieger (1971a) proposed this taxon in the introductory comments of his monograph on the Dolichomacrostomidae , and though he did not give a formal diagnosis, he mentioned enough differential characters to discern this taxon from other taxa of Dolichomacrostominae and in particular the taxon Paromalostomum Meixner, 1938 , in Ax (1951b). He also introduced the new name Austromacrostomum to contain Austromacrostomum mortenseni (Marcus, 1950) , also with eyes and paired ovaries. The female hard parts are however very different from those of the species of Cylindromacrostomum .
Two other species of Cylindromacrostomum are known (both still called members of the taxon Paromalostomum by Ax 2008): C. mediterraneum ( Ax 1955) and C. notandum (Ax 1951) . The penis stylet of the latter species has a long terminal part beyond the lamella terminal thickening, very different from what I see in my material. Only the sizes of the animal are similar ( 2–2.5 mm). The comparison with C. mediterraneum is more problematic. The animals from the type locality (Banyuls-sur-Mer, France) have a penis stylet with a short, but pointed end part that protrudes from the distal thickening, also seen in the animals from the Sea of Marmara ( Ax 1959). In the population from Venice (Lido di Alberoni) studied by Rieger (1977, Fig. 4 a) the terminal part is not pointed and barely protrudes from the distal thickening, as is also the case in the Faro population.
When considering measurements of the hard parts in the male copulatory organ, the situation becomes even more confused. An overview is given in Table I. Ax (1955; 1959) only gave the length of the accessory stylet. I have derived the measurements on the penis stylet from Ax’s figure 57 in his compilation of 2008, using the length of the accessory stylet as a ‘scale’. These measurements must thus be taken with caution!
TABLE I. Measurements of the hard parts of the male system in µm. (*own measurements in figure 57 of Ax 2008).
Do all these populations belong to the same species? One major problem is that the hard parts in the female system are insufficiently known. Ax described the middle piece as ‘kreisförmig’ in 1955 and as ‘ovoid’ in 2008. Rieger (1977, p. 215) stated that “based on the construction of female and male copulatory apparatus the Marseille individual definitely belongs to the same species as the Venice population”. Unfortunately, he did not give any information on the female system. If these two populations belong to the same species, we can assume that the (known) variation in the size of the penis stylet in these populations is, say around 145–175 µm, much longer than in the Banyuls population. The variation in the accessory stylet in the Venice-Marseille population is around 125–158 µm, also much higher than in the Banyuls population. Only the length of the terminal part of the stylet, from the proximal lamella to the end, falls in the same range in both the Venice and Banyuls populations. And, finally, the very end of the stylet is blunt in the Venice population ( Fig. 4 a in Rieger 1977), sharp in the populations A B
h
b ps e
ph
∗
t
in Ds ο
b
ο s p Βf Ff I Εf f
F mi mi mp
mp
st
st from Banyuls and the Sea of Marmara. In my opinion, the Venice-Marseille population does not belong to the same species as the one from Banyuls, the ‘true’ C. mediterraneum . Also the individual from the Sea of Marmara most probably belongs to another species, a possibility already suggested by Ax (1959).
On the Macrostomorpha Taxonomy and Phylogeny database there are micrographs and videos of two individuals, clearly representatives of the taxon Cylindromacrostomum (specimen accession codes MTP LS 546 and MTP LS 559) and with hard parts similar to those of C. mediterraneum ( sensu Rieger, 1977) . Also in these animals, the terminal part of the penis stylet is not pointed and barely protrudes from the distal thickening. They are 1–1.2 mm long, have two eyes and paired ovaries; the penis stylet is 150 and 154 mm with a terminal part of 38 and 40 mm; the accessory stylet is 133 and 162 mm (or 116–144 mm along the straight line). Shape and measures of the stylets fall exactly into the ranges given by Rieger (1977). These two individuals were found on the beach of Lignano Riviera, about 70 km from the Lido di Alberoni. There is no doubt that both populations belong to the same species. I therefore propose to consider the Venice population as a separate species, Cylindromacrostomum riegeri n.sp.. The female hard parts consist of a circular mid-piece with teeth and a diameter of 20 mm; the mouth piece is 50 mm. This also fits the drawings made by Rieger which Dr. Tyler (University of Maine, U.S.A.) kindly provided to me. C. riegeri n.sp. also occurs in the Marseille area ( Rieger 1977) and the individual from the Black Sea ( Ax 1959) might perhaps also be a member of this taxon.
The measurements of the hard parts of the copulatory organ in the population from the Ria Formosa are much greater than in C. mediterraneum from Banyuls and much lower than in C. riegeri . The distal part between the lamella and the end is slightly longer than in these two species. Only the very tip of the penis stylet resembles that of C. riegeri . In the hard parts of the female system, the mid-piece is rather similar to that of C. riegeri but the mouthpiece is only half as long. The hard parts of the female system of C. mediterraneum remain unknown. For all these reasons I cannot relate the Faro population to any of the other species of Cylindromacrostomum and consider this population to belong to a new species.

| SMNH |
Saskatchewan Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Rhabditophora |
|
Class |
|
|
SuperOrder |
Macrostomorpha |
|
Order |
|
|
Family |
|
|
SubFamily |
Dolichomacrostominae |
|
Genus |