
Aphanogmus dictynna (Waterston)
|
publication ID |
https://doi.org/10.5281/zenodo.275177 |
|
DOI |
https://doi.org/10.5281/zenodo.6218172 |
|
persistent identifier |
https://treatment.plazi.org/id/3C780612-973B-FFFE-C1BD-2B8BFBDDFD6E |
|
treatment provided by |
Plazi |
|
scientific name |
Aphanogmus dictynna (Waterston) |
| status |
|
Aphanogmus dictynna (Waterston) View in CoL
( Fig 1–10 View FIGURE 1 – 3 View FIGURE 4 – 7. 4 View FIGURES 8 – 10. 8 & 9 )
Calliceras dictynna Waterston, 1923: 116 –118.
Ceraphron dictymna (Waterston) ; Risbec, 1950: 552 (misspelling) Ceraphron dictynnus (Waterston) ; Masner, 1965: 12.
Aphanogmus dictynna (Waterston) View in CoL ; Dessart, 1971: 99; Johnson & Musetti, 2004: 14. Aphanogmus View in CoL sp. Jaramillo & Vega, 2009: 114.
Redescription. Female ( Fig 1 View FIGURE 1 – 3 ) body length: 0.82–1.00 mm. Colour: Dark brown, head almost black. Scape and pedicel yellow, remaining antennomeres mid brown. Hind coxae pale in striking contrast to the metapleura and metasoma. Hind trochanters slightly darker than coxae and all femora and tibiae mid brown, much paler than mesosoma. Terminal metasomal terga paler than preceding ones. Tarsi pale. Wings hyaline.
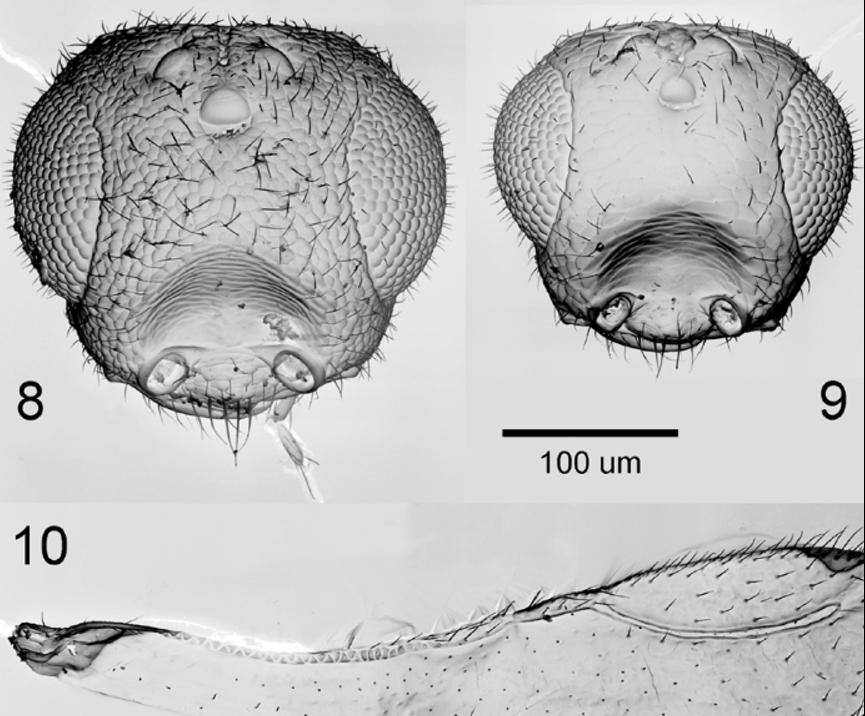
Morphology: Head globose. In dorsal view, maximum head width 1.3x head length at midpoint; maximum head width 1.5x maximum head length. Head in frontal view with antennal scrobes united into a single frontal depression. Sculpture of depression transverse, and distinct from fine reticulate sculpture of remaining frons ( Fig. 2 View FIGURE 1 – 3 ). Occeli in an equilateral triangle, lateral occeli equidistant from each other and from compound eye orbits. Anterior occelus with a small depression in front, containing a minute fovea. A shallow groove extending from stemmaticum to occiput. Antennae ( Fig. 3 View FIGURE 1 – 3 A) 10-segmented, without a distinct clava (club) unless F10, being more than 2x the length of F9, is considered as a 1-segmented clava..
Mesosoma strongly flattened laterally ( Fig. 4 View FIGURE 4 – 7. 4 ). In dorsal view appearing much narrower than head; maximum width of mesosoma 0.7x maximum head width. A single shallow median groove extending the length of the mesoscutum. Notauli and parapsidal lines absent. Mesoscutellum (scutellum) elongate-oval in dorsal view, laterally with striate carinae ( Figs 4, 5 View FIGURE 4 – 7. 4 ) characteristic of the A. hakonensis complex. Dorsellum present, unsculptured. Propodeum with a row of 8–9 deep foveae on each side. Mesosoma in lateral view ( Fig. 5 View FIGURE 4 – 7. 4 ) with fused meso- and metapleura with strong carinae fading posteriorly to an area of reticulate sculpture. Wing ( Fig. 10 View FIGURES 8 – 10. 8 & 9 ) with radial vein elongate and smoothly curving.
Metasoma in dorsal view with T1 occupying more than 0.5 of its total length; with a strong transverse carina anteriorly ( Fig. 7 View FIGURE 4 – 7. 4 ). Anterior T1 laterally with reticulate patches of sculpture each with about 6 setae, otherwise T1 completely smooth, unsculptured, with no longitudinal carinae. T3-T5 with reticulate sculpture anteriorly. S1 laterally, centrally with a reticulate patch of sculture bearing about 10 setae, ventrally more densely setose.
Male ( Figs 6 View FIGURE 4 – 7. 4 , 8–10 View FIGURES 8 – 10. 8 & 9 ) body length: 0.65–1.00 mm. Colour as female, except antennal scape and pedicel as dark as remainder of antenna. Morphology as in female except for the moniliform antennae ( Fig. 3 View FIGURE 1 – 3 b) and genitalia characters. Genitalia with the parameres ( Fig. 6 View FIGURE 4 – 7. 4 ) very strongly lobed externally, the lobes bearing long setae apically. Parameres less than half the length of the entire genital capsule, though approaching half the length in some individuals. Digiti large and conspicuous, as long as apical section of parameres, with three teeth. Aedeagus with tip blunt.
Material examined: Holotype Ƥ UGANDA: Najunga, 24.v. [19]21 H. Hargreaves coll. Presented by Imperial Bureau of Entomology. TYPE 9.121 ( NHM, on a single slide). Paratypes 4Ƥ13 same data as Holotype; 3Ƥ13 on slides, 1Ƥ card mounted. 1Ƥ13 KENYA: ex Cephalonomia stephanoderis on Hypothenemus hampei F. Infante , CIBC, ii.1982 ( NHM); 25Ƥ253, KENYA: Kisii District, Western Kenya; 00º25’S, 34º28’E, 1510m, Jarmillo & Vega, July, 2008 [ex Prorops nasuta ; specimens taken from rearing facility at ICIPE in Nairobi by first author] ICIPE, NHM USNM).
Comments. One male examined in the recently collected Kenyan material was substantially smaller (about 75% body length) than the others studied. Its small size appears to have a marked effect on head sculpture, especially of the frons (see Figs 8 & 9 View FIGURES 8 – 10. 8 & 9 ). Reticulate sculpture in the smaller individual was less rugose, with the cells much larger in relation to overall head size. Other aspects of morphology, including genitalia, suggest strongly that this is a size effect and not indicative of a different species.
Scanning electron micrographs of the fore wing base ( Fig. 10 View FIGURES 8 – 10. 8 & 9 ) reveal the structure of the basal marginal vein to be composed of successive inverted triangular elements. We have not observed this before, although it may be widespread in Ceraphronidae .
Among the described species of Aphanogmus , A. dictynna appears to be most similar morphologically to A. manilae , an extremely common and polyphagous species occurring from India to eastern Australia, and most often reared as a hyperparasitoid of pest Lepidoptera larvae. It can be separated morphologically from A. manilae by the following combination of characters: i) female antennae of A. manilae with a distinct 3- segmented clava, at least as distinguished by being dark brown in distinct contrast to the remaining antennomeres; ii) all legs pale yellow; iii) A9 of female antenna quadrate (transverse in A. dictynna ); iv) head only slightly wider than mesosoma in dorsal view (1.2x; - 1.5x wider in A. dictynna ); v) posterior meso- +metapleuron without reticulate area; vi) T1 of metasoma with many longitudinal carinae basally.
Biologically, A. manilae is most commonly associated with braconids, especially microgastrines, but has once been reared from the cocoons of a Goniozus species (Bethylidae: Bethylinae) ( Polaszek & Dessart, 1996).
Aphanogmus dictynna View in CoL appears also to be morphologically very similar to another species known from bethylids, A. goniozi Dessart (Dessart, 1988) View in CoL a species described from females only. Examination of the type series of A. goniozi View in CoL reveals that it differs from A. dictynna View in CoL in exactly the characters listed above for A. manilae View in CoL , to which A. goniozi View in CoL is apparently even more similar morphologically.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Aphanogmus dictynna (Waterston)
| Buffington, Matthew L. & Polaszek, Andrew 2009 |
Aphanogmus dictynna
| Johnson 2004: 14 |
| Dessart 1971: 99 |
Ceraphron dictymna
| Masner 1965: 12 |
| Risbec 1950: 552 |
Calliceras dictynna
| Waterston 1923: 116 |