
Bansella oregonica ( Banse, 1956 ) Banse, 1956
|
publication ID |
https://doi.org/ 10.5281/zenodo.198706 |
|
DOI |
https://doi.org/10.5281/zenodo.6200302 |
|
persistent identifier |
https://treatment.plazi.org/id/2C518784-FF8E-AC16-FF09-F99EFDE3F939 |
|
treatment provided by |
Plazi |
|
scientific name |
Bansella oregonica ( Banse, 1956 ) |
| status |
comb. nov. |
Bansella oregonica ( Banse, 1956) View in CoL , comb. nov.
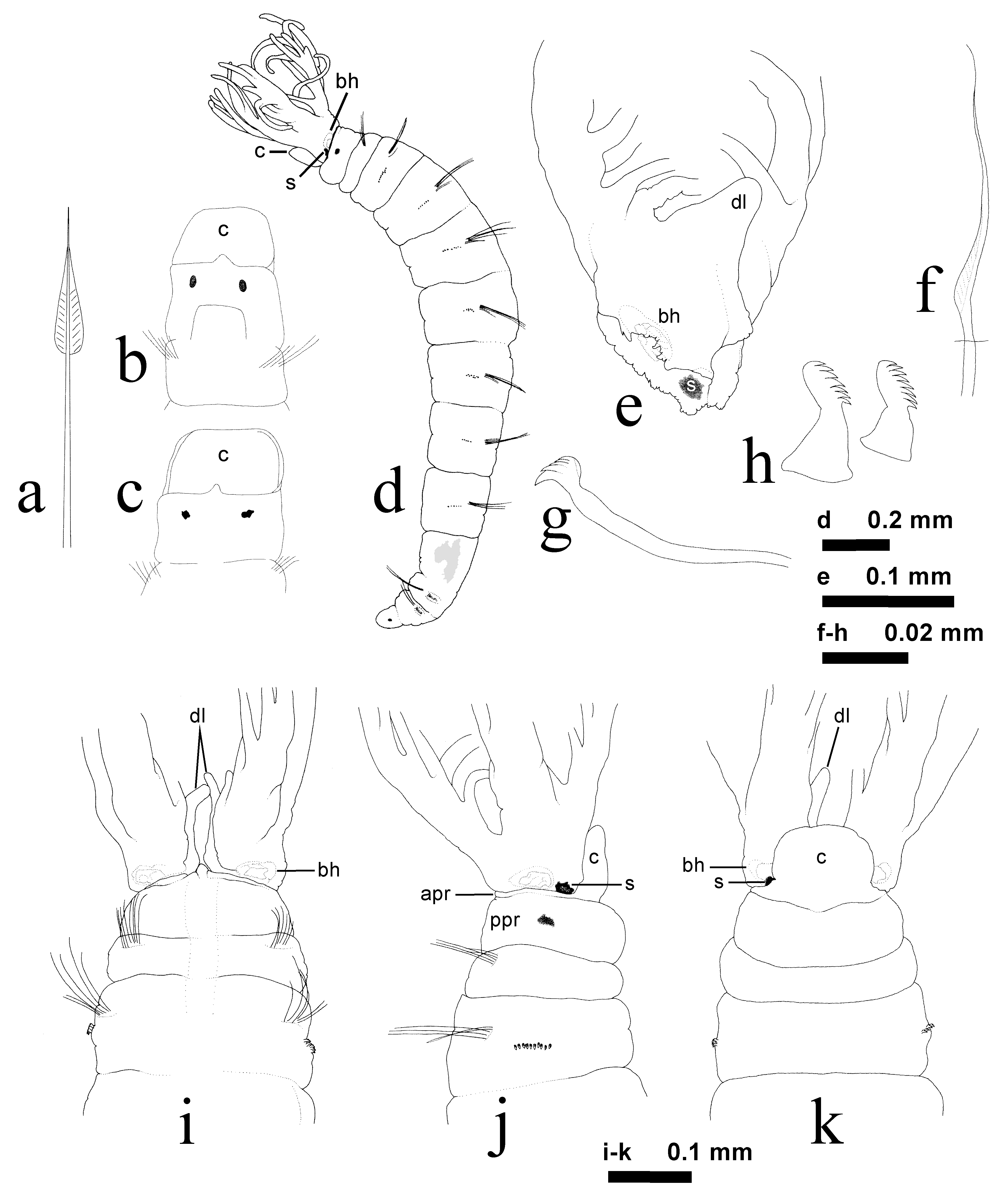
( Fig. 1 View FIGURE 1 d–k)
Fabricia dubia View in CoL not Wesenberg-Lund, 1941. — Hartman, 1951: 381, 386–387.
Fabricia sabella oregonica Banse, 1956: 426 View in CoL , 433, fig. 8.— Banse, 1963: 205, fig. 3C; Hartman, 1959: 545. Fabricia oregonica View in CoL — Banse, 1979: 874 –875; Hobson & Banse, 1981: 103, fig. 25n; Fitzhugh, 1990a: 12 –13.
Material examined. Syntypes (numerous specimens), LACM-AHF POLY 2477 [erroneously listed as AHF 3386 by Fitzhugh (1990a: 12)]. Pacific Ocean, USA, Oregon, South Bay in Coos Bay, rock washings, coll. D.H. Reish, Aug 1950.
Other material examined. Pacific Ocean, USA, Washington State, Olympic Coast National Marine Sanctuary. LACM-AHF POLY 2475 (one female, two males), Olympic Coast National Marine Sanctuary NIS-RAS sta. TB1-2, Third Beach, 47 52.586' N, 124 35.163' W, scrapings of algal turf off exposed rocks in mid-intertidal zone, coll. L.H. Harris, 23 Aug 2001. LACM-AHF POLY 2476 (3 females, 2 males), Olympic Coast National Marine Sanctuary NIS-RAS sta. NRC-3, about 0.5 miles north of Hoh River, Diamond Rock, 47 45.357' N, 124 26.843' W, river outwash of boulders, cobble, gravel between and among sea stacks, sand at bases of rhizomes of Polysiphonia sp. turf, coll. L.H. Harris, M.S. Brancato, 23 Aug 2001.
Definition. A specific hypothesis, accounting for the presence of an anterior peristomial ring with a ventral margin as a wide, distinctly rectangular lobe among observed individuals. The rectangular lobe condition originated by an unspecified mechanism(s) in a reproductively isolated population of individuals with triangular lobes, subsequent to which the new character became fixed in the population by an unspecified mechanism(s), leading to individuals observed in the present, all with anterior peristomial rings with rectangular ventral lobes.
In addition to the rectangular ventral lobe collar, members of Bansella oregonica do have a distinct combination of characters, i.e., prominent dorsal lips, absence of ventral filamentous appendages, and only narrowly hooded inferior notochaetae. Unfortunately, the phylogenetic analysis (see below) does not suggest that any of these additional characters are autapomorphies that can be accounted for in the hypothesis named B. oregonica .
Description based on Washington specimens. All specimens complete, with eight thoracic and three abdominal chaetigers. Largest specimen with branchial crown length 0.4 mm, trunk length 1.7 mm, greatest width 0.3 mm at mid-thorax; smallest specimen with crown 0.2 mm and trunk 1.4 mm. Largest specimen a complete female. Trunk slightly tapered anteriorly and posteriorly ( Fig. 1 View FIGURE 1 d). Branchial crown with three pairs of radioles; filamentous distal ends, same width as pinnules; pinnules terminate at about same height as radioles or slightly shorter. Dorsalmost radioles with 4–5 pairs of pinnules, median and ventral radioles each with four pairs. Dorsal lips erect, triangular, with distally rounded ends ( Fig. 1 View FIGURE 1 e, i, k); no associated pinnular or radiolar appendages (see Fitzhugh 2003); margins of branchial lobes ventral to dorsal lips are truncate (i.e., ventral filamentous appendages absent). Bases of ventralmost radioles with slightly raised surfaces adjacent to ventralmost pair of pinnules, but not interpreted as ventral lip-like structures (see Banse 1956: fig. 6a; Fitzhugh 1990a). Branchial hearts present. Branchial lobes not fused together mid-dorsally; lobes attached to entire peristomial margin. Females with dark brown spermatheca in ventral half of each branchial lobe ( Fig. 1 View FIGURE 1 d, e, j, k). Anterior margin of anterior peristomial ring as low ridge dorsally and laterally ( Fig. 1 View FIGURE 1 d, i–k), with narrow mid-dorsal separation; ventral ring margin as wide, rectangular lobe. Anterior peristomial ring distinctly shorter than posterior ring. Dorso-medial lobe above mouth triangular, extending slightly beyond anterior peristomial ring ( Fig. 1 View FIGURE 1 i). Anterior peristomial ring distinctly wider than long. Posterior peristomial ring about 2.5 times wider than long, and about 11 times longer than anterior ring ( Fig. 1 View FIGURE 1 d, i–k). Annulation between rings visible ventrally and laterally. Mid-dorsum of posterior peristomial ring with shallow, longitudinal depression, extending into chaetiger 2 ( Fig. 1 View FIGURE 1 i). Pair of reniform black eyes in posterior peristomial ring. Chaetiger 1 about one-half length of chaetiger 2, wider than long; chaetiger 2 about same length as posterior peristomial ring; chaetiger 3 about one-third longer than 2, with chaetiger 3–6 each about same length, slightly wider than long; chaetiger 7–8 slightly longer than preceding segments, as long as wide. chaetiger 9 about one-third length of chaetiger 8, with chaetiger 10–11 each successively shorter than 9. Pygidium about same length as chaetiger 11, posterior margin broadly tapered, rounded. Pair of round, black pygidial eyes ( Fig. 1 View FIGURE 1 d). Superior thoracic notochaetae narrowly hooded, 3–5 per fascicle. Inferior thoracic notochaetae in chaetigers 2–8 narrowly hooded ( Fig. 1 View FIGURE 1 f), distinctly shorter than superior notochaetae; 2–3 per fascicle. Abdominal neuropodia of chaetigers 9–11 with very elongate, narrowly hooded chaetae, two per fascicle. Thoracic uncini acicular, main fang slender ( Fig. 1 View FIGURE 1 g); large tooth offset from mid-line of main fang present, remaining teeth behind main fang slender and slightly decreasing in size away from fang; hood present; 8–13 uncini per fascicle in irregular single rows. Abdominal uncini with seven to eight rows of teeth in profile, three teeth per row ( Fig. 1 View FIGURE 1 h); manubrium 1.5 times longer to same length as dentate region, expanded proximally to truncate margin; uncini in chaetigers 9–11 number 16, 15, and 13, respectively. Females with oocytes in chaetigers 3–4; males with developing sperm in chaetigers 3–8, spermatids arranged in groups of more than 100, attached to central cytophore. Some specimens with light brown body wall pigmentation along inner margins of branchial lobes, peristomial rings (including collar) and chaetigers 1–2; remainder of body cream colored. Tubes not present (but see description by Fitzhugh 1990a); brooding of young not observed.
Remarks. Members of Bansella oregonica are generally similar to those of Fabricia stellaris in that both have prominent dorsal lips and the ventral lobe extension of the anterior peristomial ring. Much of the confusion in the past as to whether B. oregonica was part of the F. stellaris -subspecies group or a separate species was probably due to the incomplete comparison of individuals, as suggested by the vague descriptions presented by Hartman (1951) and Banse (1956, 1963, 1979). For instance, there is no indication that comparisons of abdominal uncini were ever made. Banse (1963) recognized the distinct differences in ventral collars in specimens belonging to F. stellaris and B. oregonica , but apparently never compared abdominal uncini. The manubrium of abdominal uncini among members of F. stellaris is 1.5 to two times longer than the dentate region ( Banse 1956: fig. 3d; Fitzhugh 1990a: fig. 1), whereas the manubrium in B. oregonica is considerably shorter ( Fig. 1 View FIGURE 1 h). Relative to members of other Fabriciinae species, there are no features uniquely shared by F. stellaris and B. oregonica . It is, however, interesting to note that in both species spermiogenesis occurs in chaetigers 3–8, which might at first suggest an exclusive sister-group relationship. As is shown in the next section, this character is not a synapomorphy. Spermiogenesis in chaetigers 3–8 also occurs among members of Pseudofabricia aberrans Cantone, 1972 (pers. obs.), several Fabriciola Friedrich, 1939 , species ( Rouse 1995b), Novafabricia labrus Fitzhugh, 1998 , and one undescribed species in the unnamed ‘Genus A’ (sensu Fitzhugh 1989; Rouse 1995b).
The present description does confirm the observations of Hartman (1951), Banse (1956, 1963), and Fitzhugh (1990a) that the inferior thoracic notochaetae are narrowly hooded in all chaetigers. Banse’s (1956: fig. 8; Fig. 1 View FIGURE 1 a) depiction of an inferior chaeta as having a somewhat “subspatulate” appearance is, however, not accurate; these chaetae have a very narrow, elongate hood ( Fig. 1 View FIGURE 1 f). The illustrations of the ventral collar lobe presented by Banse (1963; Fig. 1 View FIGURE 1 b) and Hobson & Banse (1981; Fig. 1 View FIGURE 1 c) are fairly accurate in depicting the distinct rectangular shape (see Fig. 1 View FIGURE 1 k).
None of the previous descriptions of specimens of Bansella oregonica make reference to branchial crown spermathecae ( Fig. 1 View FIGURE 1 d, e, j, k). An examination of the original LACM-AHF sample from which Banse (1956) had taken his syntypes (see Fitzhugh 1990a) revealed that spermathecae are clearly visible, albeit the specimens are in very poor condition. In addition to B. oregonica , crown spermathecae among Fabriciinae are known among members of the following genera and species: Fabricia stellaris ( Rouse 1995a: fig. 7–9), Fabriciola cri Rouse, 1996 ( Rouse 1996a: fig. 7–8, 10–12), F. l i g u ro n i s Rouse, 1993 ( Rouse 1993: fig. 31, 33), F. m i n u t a Rouse, 1996 ( Rouse 1996a: fig. 22–14, 26), Manayunkia ( Zenkevitsch 1925; Rouse 1995a: fig. 2–3, 1996a: fig. 33, 37–38; Rouse pers. comm.; not yet known in M. polaris Zenkevitsch, 1935 ), Novafabricia tenuiseta Fitzhugh, 1990 ( Rouse 1996b: fig. 1–6), and Parafabricia ventricingulata Fitzhugh, 1992 ( Rouse 1992) .
Bansella View in CoL is part of the assemblage of genera in which individuals have the following combination of features: (1) triangular dorsal lips, (2) inner, ventral margins of branchial lobes adjacent to the dorsal lips are truncate as opposed to developed as ventral filamentous appendages (see Fabriciola View in CoL , Augeneriella Banse, 1957 View in CoL ), and (3) the anterior peristomial ring collar is in the form of a ventral lobe. This group includes Fabricia View in CoL , Parafabricia Fitzhugh, 1992 View in CoL , Brifacia Fitzhugh, 1998 View in CoL , and Raficiba Fitzhugh, 2001 View in CoL , all of which are monotypic. Members of Bansella View in CoL can be readily distinguished from members of these other genera by the presence of only narrowly hooded inferior thoracic notochaetae in chaetigers 2–8, whereas pseudospatulate chaetae are present in chaetigers 3–7 in members of Fabricia View in CoL , Parafabricia View in CoL , and Brifacia View in CoL , and in chaetigers 2– 8 in members of Raficiba View in CoL . The ventral collar lobe in specimens to which Bansella View in CoL refers is also distinctive in that it is rectangular; this lobe is triangular among members of Fabricia View in CoL , Parafabricia View in CoL , and Brifacia View in CoL , and is elongate and lappet-like in Raficiba View in CoL .
| AHF |
Allan Hancock Foundation, University of Southern California |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bansella oregonica ( Banse, 1956 )
| Fitzhugh, Kirk 2010 |
Fabricia sabella oregonica
| Fitzhugh 1990: 12 |
| Hobson 1981: 103 |
| Banse 1979: 874 |
| Banse 1963: 205 |
| Hartman 1959: 545 |
| Banse 1956: 426 |
Fabricia dubia
| Hartman 1951: 381 |