
Pseudancistrus corantijniensis, Chambrier, Sophie De & Montoya-Burgos, Juan I., 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.184660 |
|
DOI |
https://doi.org/10.5281/zenodo.5622590 |
|
persistent identifier |
https://treatment.plazi.org/id/2C127E0A-3956-DF53-0393-625FFD6DF96A |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudancistrus corantijniensis |
| status |
sp. nov. |
Pseudancistrus corantijniensis View in CoL new species
Fig. 1 View FIGURE 1
Holotype: SURINAME: Sipaliwini District, Corantijn River Dr., MHNG 2672.092 (field number SU05-296). 167.16 mm SL. Corantijn River at Kaw Falls, 4°59'48.3'' N, 57°37'49.5'' W, J. I. Montoya-Burgos, R. Covain and P. Hollanda Carvalho, 5 October 2005.
Paratypes. All localities SURINAME, Sipaliwini District, Corantijn River Dr., Corantijn River, and same collectors as holotype: MHNG 2672.092, 7, 56.6-160.6 (field numbers SU05-282; SU05-292; SU05-293; SU05-297 - SU05-299); MNHN 2007-1931 (SU05-294); MNHN 2007-1932 (SU05-295); NZCS F7064 - F7065; same data as holotype. MHNG 2672.090, 1, 134.94 (SU05-309), Temery area, 4°41'10.2'' N, 57°50'03.2'' W, 5 October 2005. MHNG 2672.074, 10, 62.52-178.76 (SU05-321 - SU05-330), Wonotobo Falls, 4°22'35.3'' N, 57°57'25.1'' W, 6 October 2005. MHNG 2672.066, 1, 108.12 (SU05-355), Wonotobo Falls, 300m upstream of previous locality, 6 October 2005. MHNG 2672.056, 1, 86.0 (SU05-391), Wonotobo Falls, 4°22'35.3'' N, 57°57'25.1'' W, 7 October 2005. MHNG 2672.081, 1, 132.0 (SU05-516), Kaw Falls, 5°00'45.6'' N, 57°18'23.5'' W, 8 October 2005.
Diagnosis: Pseudancistrus corantijniensis is distinguished from non-derived Pseudancistrus and other Ancistrini by the presence of hypertrophied odontodes along the snout in males as well as in females and quite early in ontogeny. Pseudancistrus corantijniensis is diagnosed from derived Pseudancistrus on the basis of coloration: whitish spots very small, crowded and faint (especially in preserved specimens) on the snout to the posterior border of the eyes and the anterior border of the cleithrum, becoming abruptly larger (at least 1 mm in diameter even in young specimens), more spaced and more visible on the posterior part of the head ( Figs. 1 View FIGURE 1 and 2 View FIGURE 2 ). Large whitish spots continue along the body with a slight and gradual increase in size posteriorly. A single large whitish spot is generally present on each dorso-lateral plate posterior to the pelvic-fins. Pseudancistrus barbatus and P. depressus have smaller whitish spots covering the body and the spots increase slightly and gradually in size from snout to caudal peduncle rather than displaying an abrupt spot size reduction posterior to the eyes as in P. corantijniensis . In P. nigrescens , the whitish spots increase gradually in size from the snout to caudal peduncle, yet they become bigger and hazier than in the other derived Pseudancistrus . In the latter species, larger spots are often not round and can cover more than one plate.
Pseudancistrus corantijniensis can be further distinguished from other derived Guyanese Pseudancistrus by having a trapezoidal cross section in the region posterior to the head and anterior to the dorsal-fin spine versus a more round, almost semicircular cross section in other derived Pseudancistrus species. In addition, P. corantijniensis can be distinguished from P. barbatus and P. d e p re s s u s by the color and length of snout odontodes. In the new species, the odontodes are whitish and homogenous in length, with the exception of shorter ones on the tip of
Photos by S. Chraiti.
the snout, and slightly longer ones finishing on the cheek. In P. barbatus and P. depressus , the odontodes are reddish-brown (in young specimens, this color is more visible on the posterior snout odontodes) and their length increases gradually from tip of the snout to cheeks. A comparison of snout odontodes and spotting pattern on the head of the four derived Guyanese Pseudancistrus is given in Figure 3 View FIGURE 3 . Moreover, in P. corantijniensis the head is more depressed (height 7–8 times in SL; mean = 7.47, N= 23) than in P. nigrescens (height 5.3 - 6.4 times in SL; mean = 5.89, N= 12).
Description. Selected morphometrics are given in Table 1 View TABLE 1 . Pseudancistrus corantijniensis is a quite large species, largest specimen 178.8 mm in SL. Head gently sloped from snout tip to posterior part of orbit, then dorsal profile almost flat and straight to insertion of adipose-fin spine. Supraoccipital flat with slight posterior point medially. Dorsal margin of orbit higher than mesial portion of head. Iris with small dorsal flap, sometimes reaching the center of pupil. Body widest at the cleithrum with a marked and linear narrowing from cleithrum to the caudal-fin. Body dorsoventrally flattened. Dorsal profile of caudal peduncle slightly concave. Ventral surface flat. Caudal peduncle trapezoidal in cross section with ventral and dorsal surface flat, ventral surface larger.
Upper lip wide with thin papillae. Lower lip wide with thin papillae slightly larger in the central portion, posterior margin almost devoid of papillae. Only maxillary barbel present, short, sometimes bifurcated. Number of lateral plates in median series: 24. Dorsal plate series bent, flattened dorsally, forming a marked ridge from base of the dorsal-fin spine to base of caudal fin. The ridge is less marked posterior to the base of adipose-fine spine. Inframedian plate series bent in middle from the cleithrum to the insertion of pelvic-fin creating a marked keel. Ventral plates of caudal peduncle also bent, ventrally flat and forming almost a right angle on lower sides. Five rows of plates on caudal peduncle. Abdomen naked.
Numerous whitish hypertrophied odontodes along the head margin, homogenous in length excepting the anterior portion of the snout where odontodes are smaller. The few longest odontodes are on the posterior most portion of the nonevertible cheek plates. Body plates and cleithrum bear minute odontodes, which become modestly hypertrophied on the posterior margin of plates. All fin spines and rays suppor odontodes, which are moderately hypertrophied on the pectoral-fin spines, gradually larger towards the tip, in males, females and young specimens.
Dorsal-fin (I + 7) reaching adipose-fin when adpressed (mainly in young specimens). Dorsal-fin margin straight, oblique. Dorsal-fin spinlet with a concave posterior margin. Dorsal-fin lock functional. Adipose-fin with single median preadipose plate and slightly curved spine. Caudal-fin (I+14+I) margin slightly concave. Pectoral-fin (I+6) spine usually reaching the middle portion of the pelvic-fin when adpressed ventral to pelvic-fin. Pelvic-fin (I+5) spine usually reaching insertion of anal-fin spine or slightly beyond. Anal-fin (I+4) spine three fourths the length of first branched ray.
Long bicuspid teeth with a longer median lobe. 57 - 93 dentary teeth per ramus (median = 72, N = 23). 48 - 78 premaxillary teeth per ramus (median = 62, N = 23). Wide jaws, dentaries forming an oblique angle, premaxillaries almost co-linear.
Color. Ground color dark greenish-brown laterally and dorsally, becoming lighter ventrally on ventral plate series. Belly light yellow, sometimes with pinkish areas. Anterior part of head to posterior margin of orbits with many small, crowded, whitish spots; spots becoming abruptly larger and more spaced on posterior part of head and continuing onto body becoming slightly and gradually larger towards caudal peduncle. Median and supra median plate series usually with 2-3 large spots per plate anterior to pelvic-fins and one large spot per plate posterior to the pelvic-fins. Dorsal plate series with three, sometimes four spots per plate anterior to the dorsal-fin and generally two large spots per plate posterior to dorsal-fin spine. Inframedian plate series with 4-6 spots per plate anterior to the insertion of the pelvic-fin spine, generally 2-3 spots along the pelvic-fin, and generally a single spot elongated dorsoventrally in posterior plates. Ventral surface of upper lip dark greenish-brown. Dorsal-fin membrane, spine and rays with whitish oval medium size spots. Adipose-fin with one or two large whitish spots on the membrane and spine. Caudal-fin with whitish round spots centered in rays, spots slightly smaller than on the caudal peduncle. Pectoral-fin spine with whitish spots in the proximal part, fading distally; round spots centered on pectoral-fin rays. Pelvic-fin spine with or without whitish spots; when present, round spots centered on pelvic-fin rays. Anal-fin with few whitish spots or lighter areas.
Range. Known from middle and lower Corantijn River in Suriname ( Fig. 4 View FIGURE 4 )
Etymology. The name is derived from the Corantijn River, to which this species seems to be restricted.
A molecular phylogenetic approach was used to investigate species relationships among putatively derived Guyanese members of Pseudancistrus (i.e., species bearing hypertrophied odontodes along the snout). The mitochondrial D-loop sequences were obtained for 27 individuals collected in various localities and representing five species: P. barbatus (type species), P. depressus , P. nigrescens , P. n i g e r, and P. corantijniensis , n. sp. ( Table 2 View TABLE 2 ). Lasiancistrus was used as outgroup because it lacks hypertrophied odontodes along the snout and has evertible cheek plates. The sequence alignment comprised 574 positions, from which 214 where variable within the ingroup. Base composition was biased in favor of Thymine (34.8% of all bases) and Adenine (33.7% of all bases). The model of sequence evolution that fit the best our sequence data set is GTR + gamma, according to MODELTEST ( Posada and Crandall, 1998).
The maximum likelihood phylogenetic tree obtained is shown in Figure 5 View FIGURE 5 . Based on the selected outgroup ( Lasiancistrus ), P. n i g e r is distantly related to the four other members of the putatively derived Guyanese Pseudancistrus , which in turn form a monophyletic group of closely related species. This group will be referred to as the Pseudancistrus barbatus group. The type species, P. barbatus , which occurs in the Oyapock and Maroni rivers, clusters with P. depressus , for which we have samples from the Suriname and Coppename Rivers in Suriname. The closest relative to P. corantijniensis , n. sp., is P. nigrescens , collected in the Rupununi River and in the middle part of the Essequibo River in Guyana. All the inter-specific relationships are strongly supported as indicated by high bootstrap values.



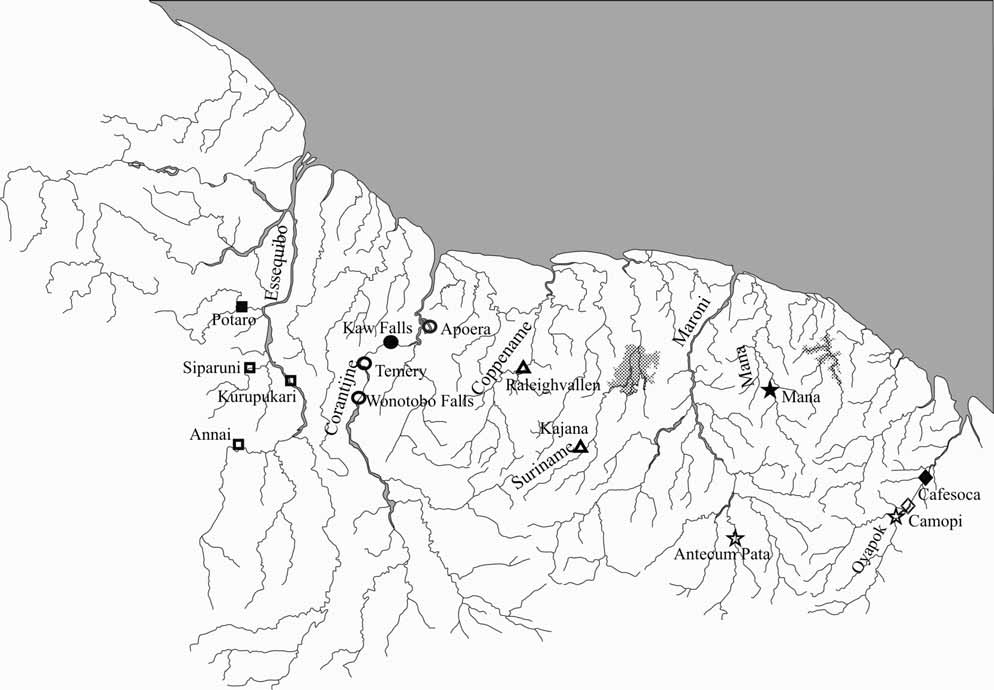
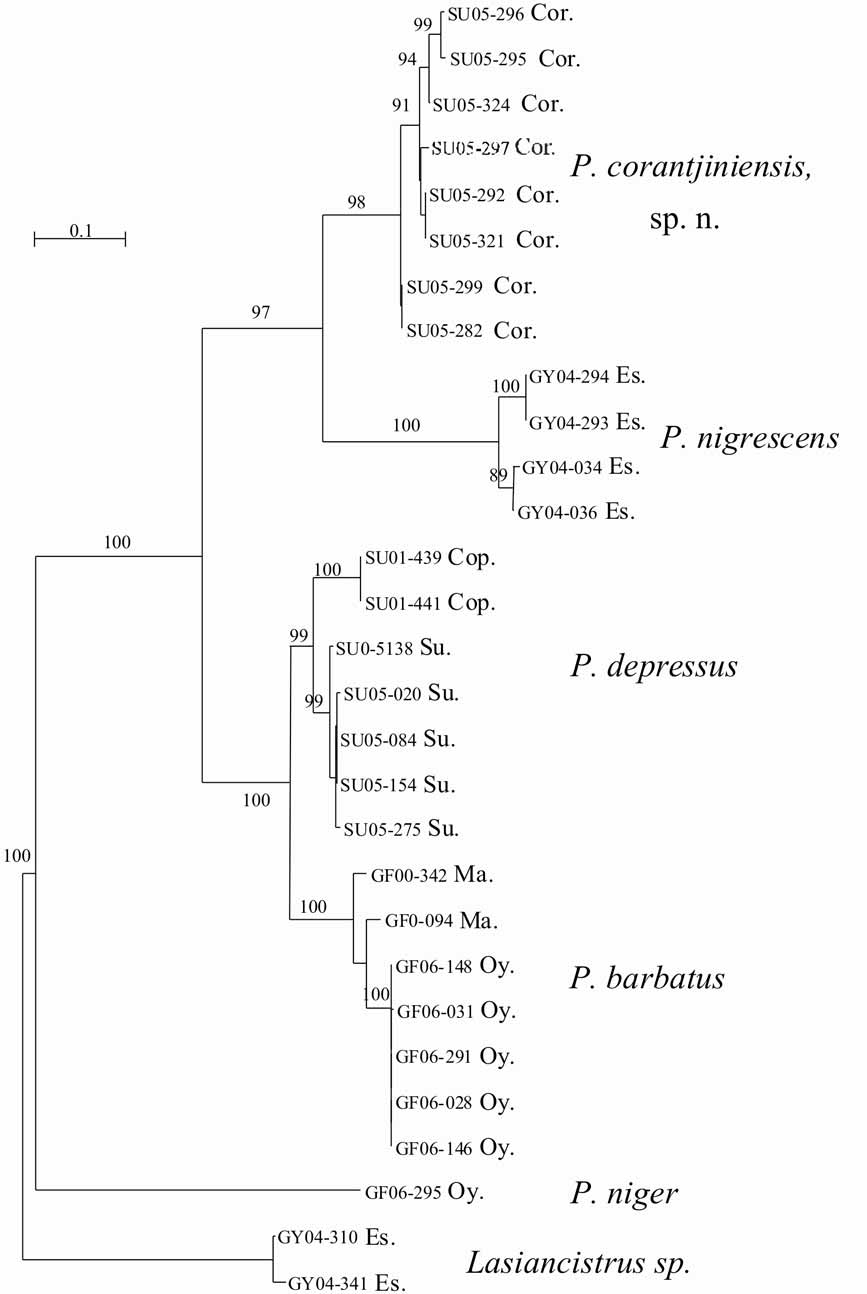
TABLE 1. Selected morphometrics of Pseudancistrus corantijniensis. Measurements 2 to 24 are percentage of Standard Length while 25 to 36 are percentage of Head Length.
| Measurements | Holotype | Average (N=23) | S.D. | Minimum | Maximum | |
|---|---|---|---|---|---|---|
| 1 | Standard Length | 160.72 | 109.1 | 37.8 | 56.6 | 178.8 |
| 2 | Predorsal Length | 41.73 | 40.74 | 1.06 | 38.78 | 42.66 |
| 3 | Head Length | 33.78 | 32.97 | 0.80 | 31.63 | 34.98 |
| 4 | Head-dorsal Length | 7.96 | 7.94 | 0.80 | 6.59 | 9.37 |
| 5 | Cleithral Width | 31.32 | 29.81 | 4.02 | 11.66 | 32.50 |
| 6 | Head-pectoral- Length | 31.07 | 27.69 | 0.67 | 26.67 | 28.97 |
| 7 | Thorax Length | 21.76 | 23.35 | 1.84 | 19.99 | 26.83 |
| 8 | Pectoral-spine Length | 36.52 | 31.47 | 2.59 | 27.16 | 36.48 |
| 9 | Abdominal Length | 26.79 | 26.34 | 1.77 | 24.28 | 30.36 |
| 10 | Pelvic-spine Length | 22.95 | 23.22 | 1.74 | 18.15 | 26.29 |
| 11 | Postanal Length | 33.97 | 32.24 | 2.05 | 24.79 | 34.91 |
| 12 | Anal-fin spine Length | 10.93 | 10.06 | 8.94 | 4.85 | 50.62 |
| 13 | Dorsal-pectoral Distance | 25.37 | 25.50 | 1.13 | 23.82 | 29.27 |
| 14 | Dorsal spine Length | 25.12 | 27.18 | 1.75 | 22.54 | 31.24 |
| 15 | Dorsal-pelvic Distance | 19.79 | 17.66 | 0.98 | 16.67 | 21.11 |
| 16 | Dorsal-fin base Length | 28.79 | 28.36 | 1.43 | 25.61 | 31.44 |
| 17 | Dorsal adipose Distance | 19.86 | 17.75 | 1.76 | 15.03 | 22.11 |
| 18 | Adipose-spine Length | 7.82 | 10.17 | 10.52 | 6.09 | 58.32 |
| 19 | Dorsal Adipose-cadal Distance | 13.6 | 12.76 | 1.05 | 10.52 | 15.26 |
| 20 | Caudal peduncle Depth | 9.98 | 9.31 | 1.59 | 6.57 | 15.44 |
| 21 | Ventral adipose-caudal Distance | 20.54 | 19.47 | 0.73 | 17.93 | 21.00 |
| 22 | Adipose-anal Distance | 21.52 | 20.53 | 1.06 | 18.17 | 22.44 |
| 23 | Dorsal-anal Distance | 14.18 | 12.72 | 0.65 | 11.08 | 13.52 |
| 24 | Pelvic-dorsal Distance | 27.2 | 25.96 | 4.25 | 8.93 | 31.20 |
| 25 | Head-eye Length | 29.46 | 35.99 | 9.10 | 28.66 | 53.00 |
| 26 | Orbit Diameter | 18.21 | 21.23 | 2.31 | 17.19 | 24.33 |
| 27 | Snout Length | 69.63 | 65.73 | 3.94 | 58.84 | 71.63 |
| 28 | Internares Width | 18.43 | 20.36 | 3.16 | 17.99 | 33.76 |
| 29 | Minimal interorbital Distance | 31.45 | 29.88 | 1.49 | 27.34 | 32.20 |
| 30 | Mouth Length | 58.87 | 58.38 | 3.97 | 51.52 | 67.43 |
| 31 | Mouth Width | 68.87 | 68.34 | 3.82 | 60.66 | 75.23 |
| 32 | Barbel Length | 10.12 | 10.53 | 14.00 | 1.86 | 61.93 |
| 33 | Dentary tooth cup Length | 21.65 | 19.48 | 4.11 | 1.51 | 22.88 |
| 34 | Premaxillary tooth cup Length | 18.67 | 17.83 | 3.41 | 4.47 | 21.31 |
| 35 | Head Depth | 39.63 | 40.66 | 1.45 | 38.38 | 44.99 |
TABLE 2. List of specimens used for the molecular phylogeny, with GenBank accession numbers.
| Species Field number | Catalog number Basin | GenBank |
|---|---|---|
| Lasiancistrus sp. GY04-310 | MHNG 2651.68 Essequibo | EF688085 View Materials |
| GY04-341 | MHNG 2650.076 Essequibo | EF688086 View Materials |
| P. n i g e r GF06-295 | MHNG 2682.037 Oyapock | EF688087 View Materials |
| P. barbatus GF06-028 | MHNG 2680.093 Oyapock | EF688106 View Materials |
| GF06-031 | MHNG 2680.093 Oyapock | EF688107 View Materials |
| GF06-146 | MHNG 281.073 Oyapock | EF688108 View Materials |
| GF06-148 | MHNG 281.073 Oyapock | EF688109 View Materials |
| GF06-291 | MHNG 2682.039 Oyapock | EF688110 View Materials |
| GF00-094 | MHNG 2653.060 Maroni | EF688111 View Materials |
| GF00-342 | MHNG 2653.063 Maroni | EF688112 View Materials |
| P. depressus SU05-020 | MHNG 2674.26 Suriname | EF688101 View Materials |
| SU05-084 | MHNG 2674.013 Suriname | EF688102 View Materials |
| SU05-138 | MHNG 2673.075 Suriname | EF688103 View Materials |
| SU05-154 | MHNG 2673.064 Suriname | EF688104 View Materials |
| SU05-275 | MHNG 2673.015 Suriname | EF688105 View Materials |
| SU01-439 | MHNG 2690.14 Coppename | EF688099 View Materials |
| SU01-441 | MHNG 2690.14 Coppename | EF688100 View Materials |
| P. nigrescens GY04-034 | MHNG 2651.022 Essequibo | EF688089 View Materials |
| GY04-036 | MHNG 2651.022 Essequibo | EF688091 View Materials |
| GY04-293 | MHNG 2650.098 Essequibo | EF688090 View Materials |
| GY04-294 | MHNG 2650.098 Essequibo | EF688088 View Materials |
| P. corantijniensis SU05-282 | MHNG 2672.092 Corantijn | EF688094 View Materials |
| SU05-292 | MHNG 2672.092 Corantijn | EF688095 View Materials |
| SU05-295 | MNHN 2007-1932 Corantijn | EF688092 View Materials |
| (Holotype) SU05-296 | MHNG 2672.092 Corantijn | EF688113 View Materials |
| SU05-297 | MHNG 2672.092 Corantijn | EF688096 View Materials |
| SU05-299 | MHNG 2672.092 Corantijn | EF688093 View Materials |
| SU05-321 | MHNG 2672.074 Corantijn | EF688097 View Materials |
| SU05-324 | MHNG 2672.074 Corantijn | EF688098 View Materials |
| Molecular Phylogeny |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |