
Capnia shasta, Nelson & Baumann, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.4758162 |
|
DOI |
https://doi.org/10.5281/zenodo.4764829 |
|
persistent identifier |
https://treatment.plazi.org/id/2B2C6D48-DD2B-FFCE-FC0D-2014FEEAFE25 |
|
treatment provided by |
Felipe |
|
scientific name |
Capnia shasta |
| status |
sp. nov. |
Capnia shasta View in CoL sp. n.
( Figs. 1-9 View Figs View Fig )
Capnia umpqua, Nelson & Baumann 1989:306 View in CoL , Figs. 177 – 180, 252, map Fig. 263 (Not Capnia umpqua Frison 1942 View in CoL )
Material examined. ♂ holotype, female allotype, and 2 male and 4 female paratypes, USA, California, Shasta Co., Sulphur Creek, Castle Crags State Park , 16 February 1985, R.W. Baumann & C.R. Nelson. The locality was checked with Google Earth internet software on 17 June 2009 and found the location just outside of the park at: N 41.15587° W 122.36285° elev. 708 m GoogleMaps . Each primary type specimen has been placed in a separate vial, in 70% ethanol. One paratype male and the allotype female were the specimens illustrated in Nelson & Baumann (1989) as C. umpqua . Of the paraptypes, one male and three females are in 70% ethanol and the terminalia of 2 males and one female are dried and on SEM stubs, including the specimens used for Figs. 1-8 View Figs (the remainders of these specimens are in 70% ethanol vials). The holotype and allotype are deposited at the Smithsonian Institution, United States National Museum , Washington, D.C. ( USNM). All paratypes are housed at the R. W. Baumann Aquatic Insect Collection, Monte L. Bean Life Science Museum, Brigham Young University, Provo, Utah ( BYUC).
Male. Body length 5.10 mm; interocular distance 0.57 mm; wings macropterous, forewing length 5.9 mm; tergum 9 divided along one-half its length by medial membranous area, anterior margin heavily sclerotized. Well-developed tergal knobs present along inner margin of the divided tergum nine ( Figs.1-4 View Figs ), no tergal knobs on segment eight; epiproct broad, length 285 m, epiproct width 132 m, measured in dorsal view ( Figs. 1, 3 View Figs ) with only a single process well-developed ( Figs. 2, 4 View Figs ); apex of epiproct unforked ( Figs. 1, 3, 5 View Figs ).
Female. Body length 6.48 mm; interocular distance 0.62 mm; wings macropterous, forewing length 6.80 mm; subgenital plate triangular with apex directed anteriorly ( Figs. 8 View Figs , 9 View Fig ); subgenital plate width 0.46 mm on hind margin; hind margin straight with margin thickened and not overlapping sternite 9; subgenital plate with paired lateral light thinnings of sclerotization near distal corners, immediately anterior to the hind margin, darker lines (probably internal) running much of the length of subgenital plate near medial line ( Figs. 8-9 View Figs View Fig ).
Etymology. This species is named for prominent Mount Shasta of the Cascade Range in northern California as a noun, without gender, in apposition. It shares this name with a common, somewhat generic brand of soda water common in the western United States.
Diagnosis. Capnia shasta is unique in that it is the only member of the Californica Group with tergal knobs only on segment 9 and no distinctively divided process on the epiproct. It lacks the distinct upper process of the epiproct found in the Ventura Subgroup. It differs from the Californica Subgroup in lacking knobs on tergum 8. Phylogenetic relationships of C. shasta with all known members of the Californica Group are summarized in Fig. 10 View Fig .
Discussion. Nelson & Baumann (1989) predicted that the area in California between Santa Barbara and Carmel would probably be a site for a new species or two yet to be discovered in the Californica Group. To this date, no one has reported any new records of Capnia species from this area but in this paper we describe C. shasta , from a more northerly thought-to-be-well-collected and central portion of the overall group range. The challenge remains to find coastal new members of the group in northern California.
Key to males. All males of the previously known species in the Californica Group except true C. umpqua can be identified using the key (p. 304) of Nelson & Baumann (1989). The following key is a modification of the 1989 key written to accommodate both true C. umpqua and C. shasta . Couplet numbering follows that key, with couplet 8 added and figures from various sources noted.
7(4) Terga 5 and 6 each bearing paired knobs (Figs. 69 and 70 of Nelson & Baumann 1989) …………………………………… jewetti Frison View in CoL
7’ Terga 8 or 9 bearing paired knobs ( Figs. 1-4 and 7 View Figs in this publication and Figs. 177-178 of Nelson & Baumann 1989) ………………….... 8
8(7) Terga 8 bearing paired knobs ( Figs. 1-2 View Figs in Baumann & Stewart 2009) ……. umpqua Frison View in CoL
8’ Terga 9 bearing paired knobs ( Figs. 1-4 View Figs in this publication; Figs. 177-180 in Nelson & Baumann 1989; and Figs. 3-4 View Figs in Nelson 2004 …………………………….. shasta View in CoL , new species
Phylogeny. A purported phylogeny for the entire Californica Group was produced in ( Nelson 2004). In the 2004 paper, the species illustrated (and used for the phylogenetic analysis) as C. umpqua was the species we herein describe as C. shasta . Table 1 View Table 1 summarizes the morphological characters used in the 2004 paper with addition of character states observed from the holotype of C. umpqua , a minor reinterpretation of Character 14, and a correction of species name for the Sulphur Creek, Castle Crags State Park specimens ( C. umpqua of Nelson 2004) as C. shasta . We ran a branch and bound analysis using this corrected matrix ( Table 1 View Table 1 ). This analysis yielded six equally parsimonious trees. In all of these trees, the topology conformed largely to that of the tree in Nelson (2004) except that now C. nana falls into a polytomy with C. shasta and the two subgroups ( Fig. 10 View Fig ). Still C. shasta is basal to C. jewetti . In three of these trees C. shasta is immediately basal to C. jewetti and in the remaining three it is also basal to C. regilla . Thus in the consensus of the six trees it appears as a polytomy with the two major clades in the Californica Group ( Fig. 10 View Fig ) and C. nana . The more important question then becomes, “Where does true C. umpqua fit on the tree?” In five of the six most parsimonious trees, C. umpqua fits in a pectinate fashion up the tree from C. jewetti and down from C. ophiona . In the sixth tree C. umpqua is still up tree from C. jewetti but in a polytomy with C. ophiona and ( C. californica + C. quadrituberosa ).
Zoogeography. With the consensus cladogram of purported relationships ( Fig. 10 View Fig ) and the overall distribution of the group’s species in hand ( Nelson 2004) one is tempted to hypothesize the origins, dispersal, and speciation sequence for members of the group. The zoogeography for the group as previously presented ( Nelson 2004) stands. The Californica Group, taken as a whole ( Nelson 2004, Fig. 3 View Figs ) divides into two sister taxa basally (Ventura Subgroup) and (Californica Subgroup + C. umpqua + C. jewetti ). But our new species comes out in a polytomy with these two subgroups (and C. nana ) and thus leads to no strong zoogeographic hypotheses.
Capnia shasta is currently known from a single locality on a single date. It is currently known only from far down the flanks of Mount Shasta in a small tributary of the Sacramento River at the northern head of California’s Central Valley. We suggest that further careful winter collections of many sites in the area are warranted. We examined a range of specimens of C. umpqua from throughout its broad Oregon – southern California range. The shape of the epiproct in these specimens varied widely in relative width both within series from a single collection and across its range. Using this information we tentatively propose that C. shasta , C. jewetti , and C. ophiona arose as independent peripheral isolates from the widespread C. umpqua -like ancestor. This speculative hypothesis could best be tested using fast evolving sequences of DNA. We have little hope that further morphological study of this question will resolve these polytomies, and hope that the sequences will.

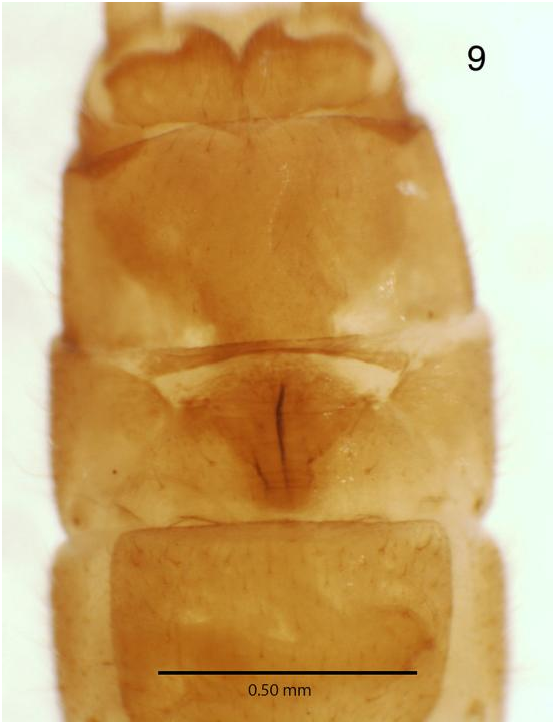
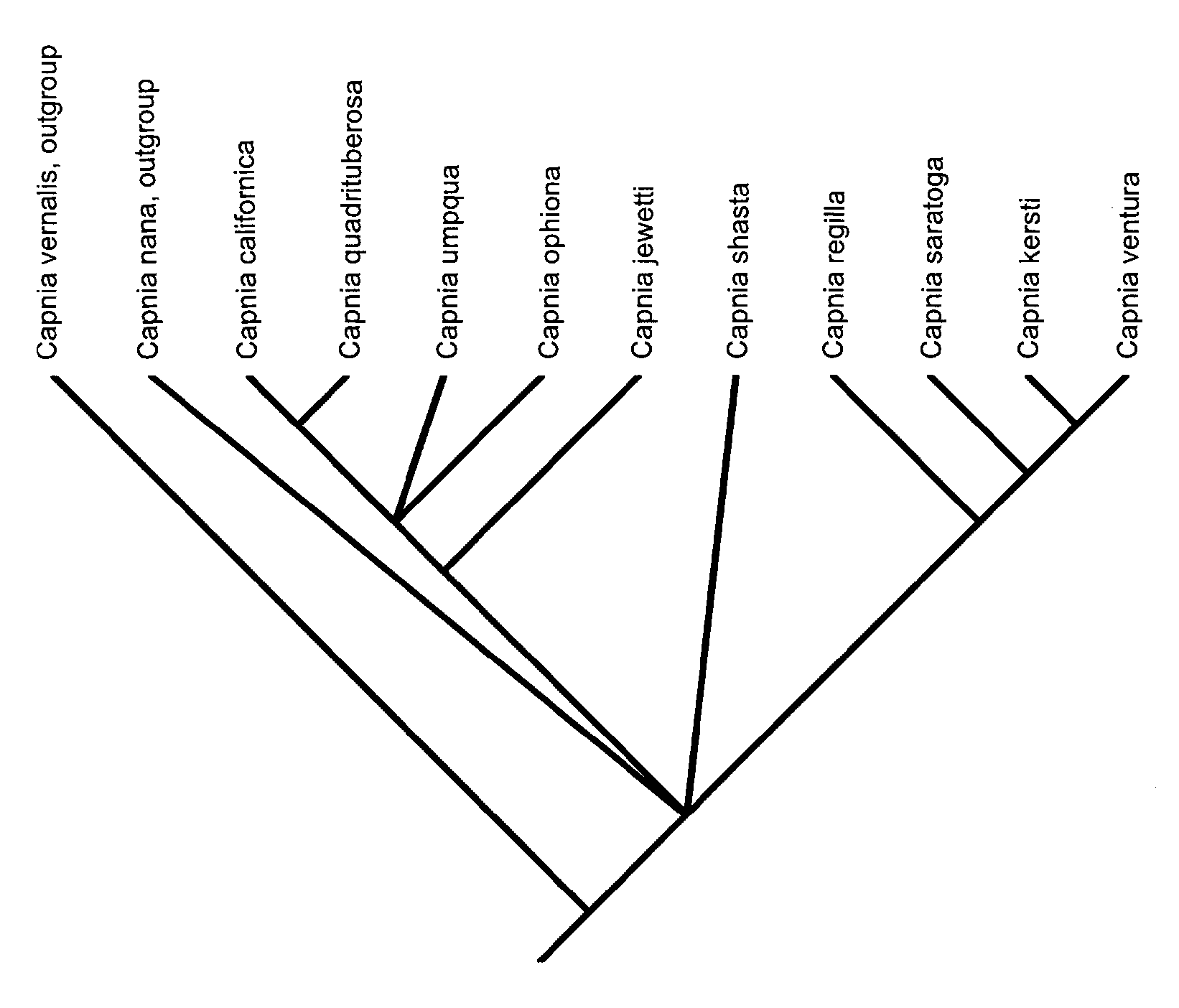
Table 1. Matrix of characters used in the parsimony analysis of the Californica Group, with Capnia vernalis and C. nana as outgroups. Character states updated from Nelson (2004), with the correction of data related to C. umpqua and addition of data for C. shasta. State of character 14 is corrected for C. regilla from 0 in Nelson (2004) to 1. All character state texts given are for the “1” state. Question marks (?) indicate missing data. Characters: 1. Knobs present on tergite 8; 2. Knobs present on tergite 9; 3. Lower process of epiproct forked. 4. Female subgenital plate overlapping posterior margin of sternum 8. 5. Upper process of epiproct absent (or very greatly reduced) apically. 6. Upper process of epiproct divided in lateral view. 7. Declivity between processes of epiproct vertical. 8. Lower process of epiproct longer than upper process. 9. Lower process of epiproct much longer than upper process. 10. Knobs present on tergites 5 and 6. 11. Both processes of epiproct of about the same length. 12. Lower process of epiproct widely forked in dorsal view. 13. Declivity between processes of epiproct medially carinate. 14. Declivity between upper and lower processes of epiproct with an extra medial process. 15. Knobs on tergite 8 angulate. 16. Notch between knobs of tergite 8 broad in dorsal view. 17. Notch between upper and lower processes deeply incised in lateral view. 18. Lower process of epiproct narrow in dorsal view. 19. Upper and lower processes of epiproct forming a 90 degree angle in lateral view. 20. Lower process of epiproct with lower surface bent at mid length in lateral view. 21. Lower process of epiproct constricted near apex in dorsal view. 22. Lower process of epiproct evenly tapered, not inflated at midlength in dorsal view.
| Taxon | Characters | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 22 | ||
| Capnia vernalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia nana | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia californica | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia quadrituberosa | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia ophiona | 1 | 1 | 1 | ? | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia jewetti | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia regilla | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia saratoga | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 0 |
| Capnia kersti | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| Capnia ventura | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 |
| Capnia umpqua , holotype | 1 | 1 | 1 | ? | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Capnia shasta , n. sp. | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Capnia shasta
| Nelson, C. Riley & Baumann, Richard W. 2009 |
Capnia umpqua
| Nelson, C. R. & R. W. Baumann 1989: 306 |