
Hemicyclops ventriplanus, Kim, 2000
|
publication ID |
https://doi.org/10.1080/002229300299543 |
|
persistent identifier |
https://treatment.plazi.org/id/291F7827-E95B-092E-FE82-FE6DFDF63547 |
|
treatment provided by |
Felipe |
|
scientific name |
Hemicyclops ventriplanus |
| status |
sp. nov. |
Hemicyclops ventriplanus n. sp.
( Figures 1 View FIG ±3)
Material examined. Numerous specimens, including copepodids I± V, found in the washings of the thalassinidean crustacean, Upogebia major (De Haan) , collected from the intertidal mud ¯at at Jakyak-do Island, o Inchon, on 28 September 1996. Holotype (ovigerous m), allotype (l), and 80 paratypes ( 50 mm, 30 ll) have been deposited in the US National Museum of Natural History , Smithsonian Institution. Other specimens are kept in the collection of the author .
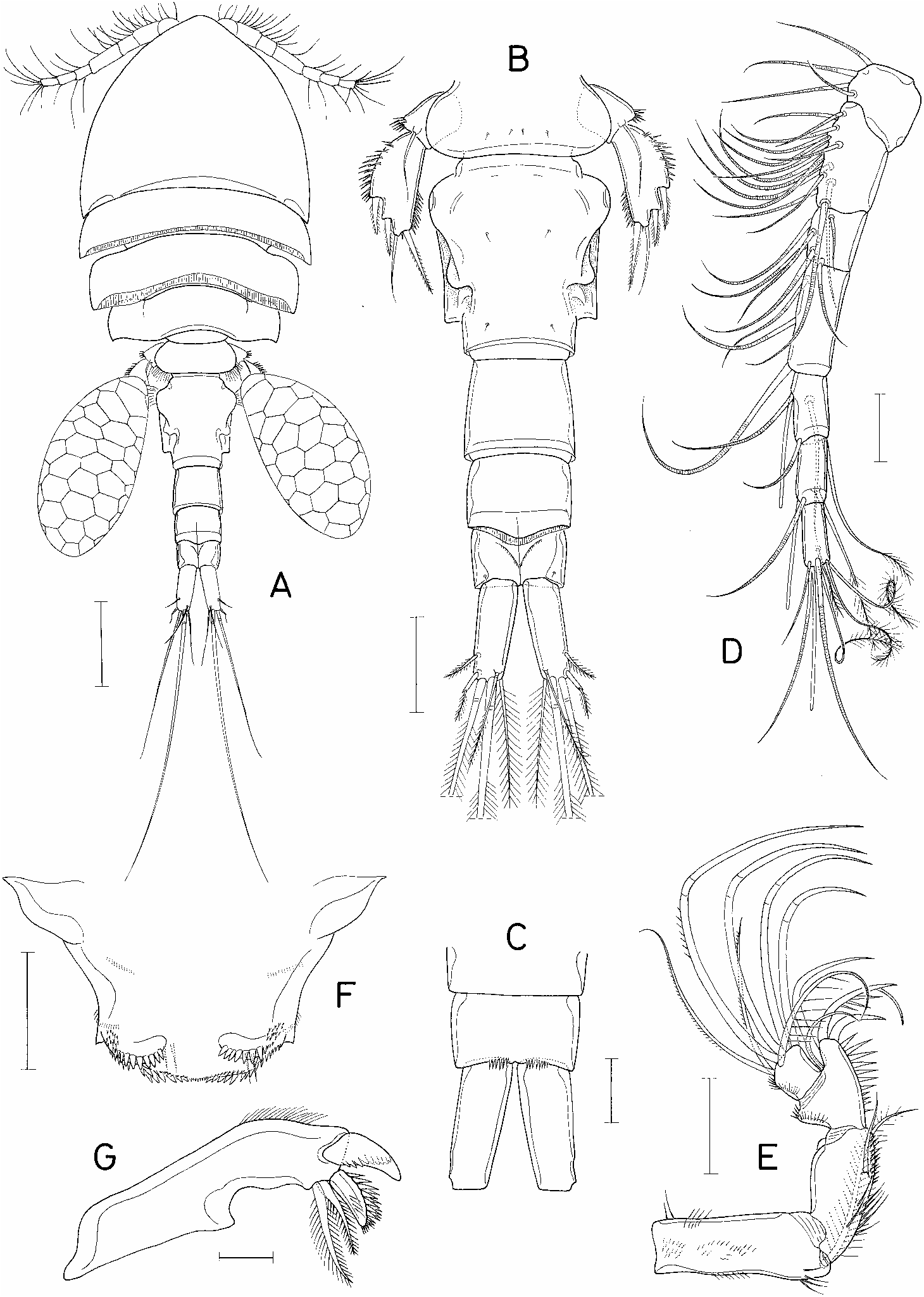
Female. Body (®gure 1A) cyclopiform. Selected specimen 1.39 mm long. Greatest width 0.55 mm. Prosome composed of cephalothorax and three pedigerous somites. Rostral area of cephalosome produced anteriorly. Prosomal somites with well developed epimera. Second and third pedigerous somites with posterior margin of tergite fringed with membrane. Fourth pedigerous somite with pointed posterior corners. Urosome (®gure 1B) ®ve-segmented. Fifth pedigerous somite 194 mm wide, with round lateral margins. Genital double-somite ventrally ¯at, dorsally complex as in ®gure 1B, 212Ö 188 mm (1.13:1). Genital area located laterally near anterior portion of somite at area of greatest width. Three abdominal somites 103 Ö117, 74 Ö109, and 62Ö 101 mm, respectively. Posterodorsal border of second abdominal somite convex, fringed with membrane. Anal somite with spinules on medial side of posteroventra l border (®gure 1C). Caudal ramus 102Ö 40 mm (2.55:1), with nearly parallel lateral margins and six setae. Outermost seta divided into two parts: thick basal and plumose distal parts. Longest inner median terminal seta 653 mm. Next longest, outer median terminal seta 359 mm. Egg sac oval (®gure 1A), 441Ö 223 mm.
Rostrum incorporated with ventral surface of cephalothorax. Antennule (®gure 1D) seven-segmented, 363 mm long, with armature formula 4, 15, 6, 3, 4 +1 aesthetasc, 2 +1 aesthetasc, and 7 +1 aesthetasc. Antenna (®gure 1E) foursegmented, with armature formula 1, 1, 4, 7. Third segment with distinct inner distal protuberance and spinules on inner margin. Terminal segment slightly wider than long.
Labrum variously ornamented with spinules and setules as ®gure 1F. Mandible (®gure 1G) armed terminally with four elements: one dorsal, plate-like element bearing denticles, another plate-like element bearing spinules on dorsal side, and two ventral plumose setae. Paragnath a lobe bearing distal process and a posterior row of spinules (®gure 2A). Postoral region armed as in ®gure 2A. Maxillule (®gure 2B) distally bilobed, with ®ve setae on larger lobe and three setae on smaller lobe. Maxilla (®gure 2C) two-segmented. Basal segment with a row of setules basally and two distal, barbed setae, one of them accompanied basally with one setule. Maxilliped (®gure 2D) four-segmented, with formula of armature: 2, 2, 0, 3 +II. First and second segments each with row of spinules in addition to setae. Inner margin of second segment produced near base of setae. Terminal spine on terminal segment with one basal, terminally-forked seta and six or seven setules on outer side of proximal half. Inner spine enlarged, with several sti spinules on both margins.
Legs 1±3 (®gure 2E±G) and leg 4 (®gure 3A) with three-segmente d rami. Formula of armature of these legs as follows:
Median portion of posterior margin of basis of legs 1±4 spinulated. Posterior margin of intercoxal plate of leg 1 with hairs, but those of legs 2±4 with spinules. Outer margin of basis in leg 1 with setae, but those in legs 2±4 with spinules. Inner spine of basis of leg 1 slender and longer than ®rst endopodal segment. Outer margin of endopod of all biramous legs with hairs. Outer margin of ®rst exopodal segment smooth but those of legs 2±4 with spinules.
Leg 5 (®gure 3B) two-segmented. Basal segment with one seta, and spinules near base of seta. Distal segment 103Ö 51 mm (ratio 2.02:1), with three spines, one seta, and spinules on outer margin and on distal half of inner margin. Leg 6 not observable, concealed from dorsal and ventral views of body.
Spermatophor e 120Ö 62 mm, a curved sac as ®gure 3C.
Male. Body (®gure 3D) resembling that of female. Length 1.38 mm. Urosome (®gure 3E) six-segmented. Fifth pedigerous somite 212 mm wide, narrowed posteriorly. Genital somite nearly hexagonal, 188Ö 218 mm. Four abdominal somites 88 Ö126, 91 Ö106, 62 Ö100, and 52Ö 95 mm, respectively. Caudal ramus 91Ö 38 mm (2.39:1).
Antennule, antenna, labrum, mandible, paragnaths, and maxillule identical to those of female. Maxilla (®gure 3F) sexually dimorphic; second segment terminated in a claw-like, blunt process armed distally with minute spinules. Maxilliped (®gure 3G) composed of three segments and terminal claw. First segment with one long plumose seta. Second segment greatly expanded proximally and narrowed distally, with one row of minute spinules, two rows of larger spinules and distantly separated, two small setae on inner surface. Third segment unarmed. Claw weakly curved, proximally with one seta and one spinule.
Leg 1 (®gure 3H) without inner spine on posterior margin of basis. Legs 2±4 as in female. Basal segment of leg 5 completely incorporated into ®fth pedigerous somite. Free segment identical in shape to that of female. Leg 6 represented by two small spines on genital ¯ap.
Etymology. The speci®c name ventriplanus is a combination of the Latin venter (= the belly) and planus (= ¯at). It alludes to the ¯at ventral surface of the female genital double-somite.
Remarks. The new species bears the following diagnostic features: (1) eight (neither six nor nine) elements on the third exopodal segment of leg 4; (2) two (rather than one) setae on the second endopodal segment of leg 4; (3) four (rather than ®ve) setae on the ®rst segment of the antennule; (4) ®ve- (rather than six-) segmented urosome in female; and (5) four (rather than three) elements on mandible.
According to Humes (1995), 36 species are recognized in the genus Hemicyclops . The above combination of characters is shared with more than half of its congeners. Nine of them have, as in the new species, a caudal ramus length to width ratio ranging between 2 and 3: H. acanthosquillae Humes, 1965 ; H. axiophilus Humes, 1965 ; H. bacescui (SË erban, 1956); H. biāgellatus Humes, 1965 ; H. gom soensis Ho and Kim, 1992 ; H. kombensis Humes, 1965 ; H. mortoni Boxshall and Humes, 1988 ; H. thalassius Vervoort and Ramirez, 1966 ; H. visendus Humes, Cressey and Gooding, 1958 .
Of these nine species, H. thalassius shows no sexual dimorphism in leg 1, i.e., its male carries an inner spine on the basis and therefore cannot be confused with the new species. The male of both H. bacescui and H. mortoni are not known, but the remaining six species have in common males that possess no spine on the basis of leg 1 and only a single seta on the syncoxa of the maxilliped. Morphological features which distinguish H. ventriplanus from these eight species are given below.
In H. acanthosquillae , H. biāgellatus , and H. kombensis , all of them described from Madagascar, the genital double-somite consists of an anterior expansion (although weak in H. kombensis ) and the posterior part is long and narrow with parallel lateral margins. Furthermore, the inner distal angle of the antenna of H. acanthosquillae is prominently projected so that it extends beyond the end of the fourth segment. In H. biāgellatus , the inner spine on the basis of female leg 1 is very long, extending beyond the distal border of the endopodal second segment. The free segment of leg 5 in H. kombensis is characteristic because it is very narrow proximally but strongly expanded distally.
The genital double-somite of H. axiophilus has two prominent, anterior and posterior, expansions. In H. bacescui the genital double-somite has a dorsal surface suture ( Stock, 1959), but is otherwise cylindrical. Hemicyclops visendus also has a simple, quadrangular genital double-somite and a stocky free segment of leg 5.
The male leg 6 of H. gomsoensis is represented by two spines. In this and other morphological aspects, this other Korean species is very closely related to the new species. The signi®cant di erences between the two species can be found in the shape of the female genital double-somite and of the spermatophore. The genital doublesomite of H. gomsoensis shows a distinct constriction in the middle and a pair of dorsal ¯aps, while in the new species neither constrictions nor ¯aps are present. The spermatophore of H. gomsoensis is long and slender, whereas that of the new species is vesicular.
The genital double-somite of H. mortoni also bears a constriction in the middle and a pair of prominent, lobate projections near each of the posterior corners. The free segment of leg 5 is stockier (ratio 1.61:1) than that of the new species (ratio 2.02:1). The maxilla of H. mortoni is peculiar, because its main terminal element of the second segment is a simple process bearing two cusps (®gure 1H in Boxshall and Humes, 1987). This main terminal element in the new species is a thick, bifurcate spine (or process) bearing three cusps. The third endopodal segment of leg 4 is armed with four spines and one seta (IV, 1) in H. mortoni , compared with three spines and two setae (III, 2) in the new species.
Hemicyclops japonicus , a species discovered from a dredged area in Tokyo Bay, Japan ( Itoh and Nishida, 1993), is distinguishable from the new species by its broader caudal rami (ratio 1.7:1) and the di erent structure of male leg 6. Otherwise they are very similar to each other. In particular, the Japanese species bears a pair of lateral processes (Itoh and Nishida called them`hooks’) in the posterior third of the genital double-somite resembling the posterolateral angles of the new species. The oral appendages and leg armature are also very alike in the two species. Itoh and Nishida (1993) considered H. axiophilus as the species most closely related to H. japonicus . It is interesting that both H. axiophilus and H. ventriplanus are associated with thalassinidean hosts. This fact further suggests that H. japonicus may well turn out to be an associate of a thalassinidean.
| V |
Royal British Columbia Museum - Herbarium |
| US |
University of Stellenbosch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hemicyclops ventriplanus
| Kim, Il-Hoi 2000 |
H . ventriplanus
| Kim 2000 |
Hemicyclops japonicus
| Itoh & Nishida 1993 |
H . japonicus
| Itoh & Nishida 1993 |
H . japonicus
| Itoh & Nishida 1993 |
H . axiophilus
| Humes 1965 |
H . axiophilus
| Humes 1965 |