
Gekko gecko
|
publication ID |
https://doi.org/ 10.5281/zenodo.278393 |
|
DOI |
https://doi.org/10.5281/zenodo.6188384 |
|
persistent identifier |
https://treatment.plazi.org/id/19358A1A-8B01-FFA4-8980-FAB8FD6EF133 |
|
treatment provided by |
Plazi |
|
scientific name |
Gekko gecko |
| status |
|
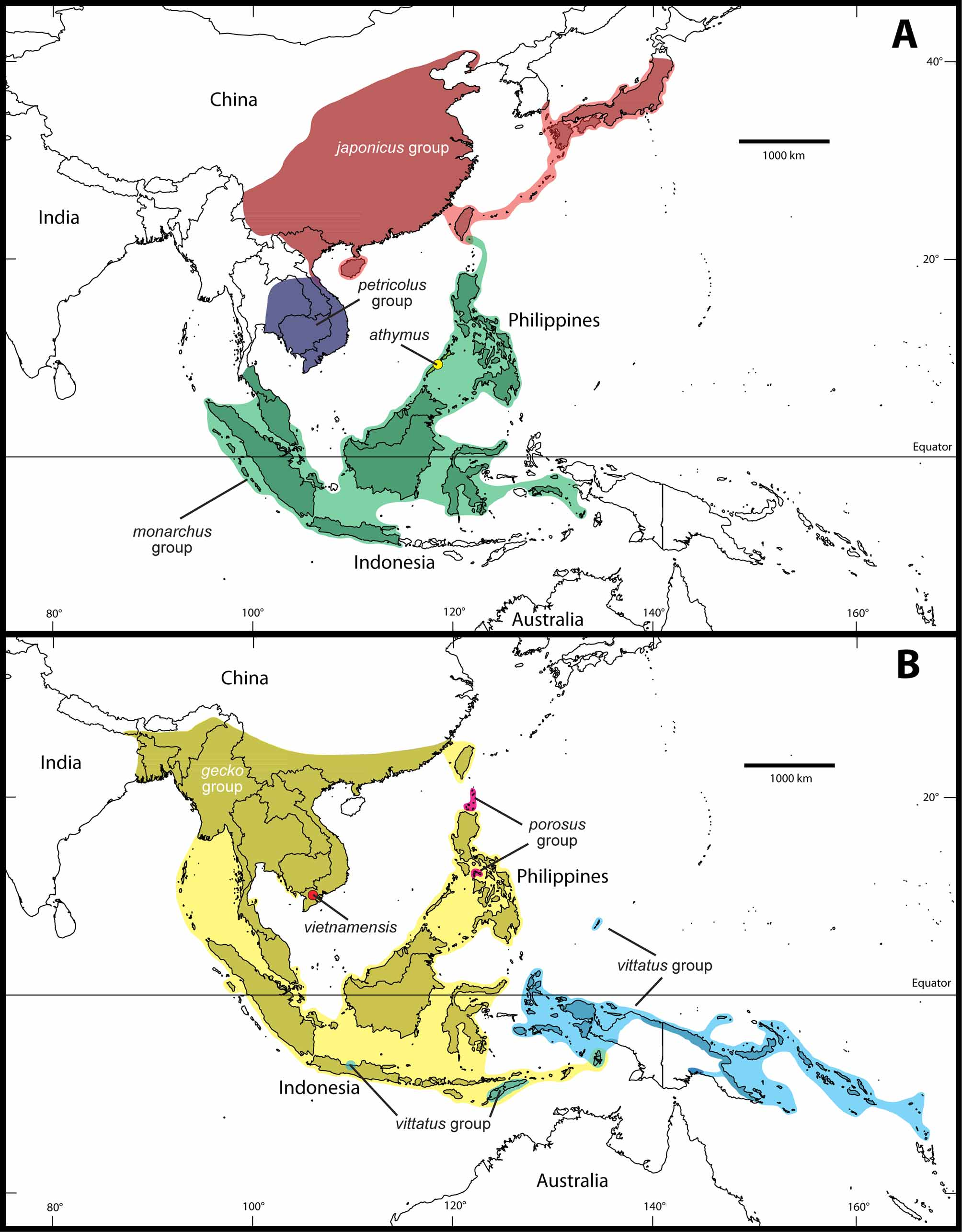
Gekko gecko group
( Figure 2 View FIGURE 2. A )
Species. G. albofasciolatus Günther, 1867 ; G. gecko ( Linnaeus, 1758) ( Figure 2A View FIGURE 2. A ); G. nutaphandi Bauer, Sumontha & Pauwels, 2008 ; G. siamensis Grossmann & Ulber, 1990 ( Figure 2 View FIGURE 2. A B); G. smithii Gray, 1842 ( Figure 2 View FIGURE 2. A C); G. verreauxi Tytler, 1864 ( Figure 2 View FIGURE 2. A D); Gekko sp. ”Tioman“; Gekko sp. ”Sulawesi“; Gekko sp. ”Togian.”
Characters. 150.0–191.0 mm SVL; nares, except for G. verreauxi , not in contact with rostral; nasals 3–6; postmentals relatively slow (largest in G. siamensis ), dorsal tubercle rows 10–19; precloacal pores 10–16; postcloacal tubercles 2–4 (rarely single); webbing between fingers and toes lacking; tubercles present on fore and hind limbs; lateral fold without tubercles; subcaudals enlarged, in two parallel rows; iris yellow, green, blue or brick red; Yshaped head pattern usually discernible; light (white), more or less transversally arranged, symmetrical dorsal and lateral blotches.
Distribution. From India and Nepal to China, southwards to Indonesia (Indoaustralian Archipelago) ( Figure 3 View FIGURE 3 B); G. albofasciolatus is endemic to Borneo; G. verreauxi is endemic to the Andaman Islands; Gekko sp. "Tioman" is endemic to Tioman Island; Gekko sp. "Sulawesi" is endemic to Sulawesi; Gekko sp. "Togian" is endemic to Togian and Batudaka. Allochthonous populations of Gekko gecko exist in the Caribbean, in Belize, on Hawaii and in Florida ( Kraus 2009).
Remarks. In the original description of Gecko albo-fasciolatus (= Gekko albofasciolatus ), Günther (1867) only provided a vague type locality Polynesia with a question mark. Smith (1935) subsequently stated “ type loc. unknown, probably Malay Archipelago.“ Later, Wermuth (1965) wrote „Polynesia“ as the type locality with the addition (fide Smith 1935), that the species probably occurs in the Indo-Australian Archipelago. According to Günther (1872), G. s m i t h i i occurs in the north of Borneo (Labuan, now Sabah) and G. albofasciolatus in the South (Banjermassin, Martapoura). We herein restrict the type locality of G. albofasciolatus to Banjermassin (= Banjarmasin), Kalimantan, Indonesia. While commenting on G. albofasciolatus, Günther (1872: 589) stated that he received three specimens from Dr. Bleeker from Borneo under different names ( Platydactylus pentonopus , Platydactylus borneensis , and Hemidactylus zosterophorus ). From Günther’s (1872) footnotes it is obvious that he allocated all of Bleeker’s specimens to G. albofasciolatus . Of Bleeker’s species, only Platydactylus borneensis had been recorded from Borneo. The names P. borneënsis (= P. borneensis ) and H. zosterophorus were introduced by Bleeker (1857) without formal species descriptions (see also Bleeker 1860). The type loaclity of Platydactylus borneensis (non Pentadactylus borneensis Günther, 1864 = Aeluroscalabotes felinus Günther, 1864 ; non Tarentola borneensis Gray, 1845 = Tarentola delalandii Duméril & Bibron, 1836 ) is “Bandjermasin” and the type locality of Hemidactylus zosterophorus is “Padang (ook op Nias)”. In his critical review of Bleeker’s type specimens, Boulenger (1887) did not consider the taxa Platydactylus borneensis and Hemidactylus zosterophorus . According to Bauer (1994), both names are species inquirenda and Kluge (2001) listed them as nomina nuda (see also Rösler 2000). Therefore, the names Platydactylus borneensis Bleeker, 1857 and Hemidactylus zosterophorus Bleeker, 1857 , which are according to Günther’s (1872) statements younger, subjective synonyms of Gekko albofasciolatus Günther, 1867 , are not available. The same concerns Günther’s (1872) name Platydactylus pentonopus . Boulenger (1885) synonymized G. albofasciolatus with G. s m i t h i i (see also Wermuth 1965; Kluge 1991, 1993; Bauer 1994; Rösler 2000). De Rooij (1915) obviously also followed Boulenger (1885), because she listed the distribution of G. albofasciolatus based on Günther (1872) under the species G. smithii . Recently, Kluge (2001) revalidated G. albofasciolatus at the specific rank, but Malkmus et al. (2002) and Das (2004) did not consider G. albofasciolatus as part of the Bornean herpetofauna. According to its original description ( Günther 1867), G. albofasciolatus differs from G. s m i t h i i by a different coloration of the dorsum (reddish-olive, marbled with grayish in G. albofasciolatus ) and the venter (lower parts uniform whitish in G. albofasciolatus ). Both species correspond well with respect to size and characteristic scalation (e.g., nares not in contact with rostral; postmentals relatively short; number of supralabials and sublabials; dorsal tubercle rows; precloacal pores) as well as in head and back pattern. A distinguishing feature at present is the number of ventrals (26 in G. albofasciolatus versus 29–39 in G. smithii ). However, further specimen-based studies are needed to confirm whether G. albofasciolatus in fact deserves specific recognition.
Concerning G. s m i t h i i, Das (2004, 2007) depicted specimens from Borneo, which did not show a Y-shaped head pattern, as is characteristic for specimens from Thailand and Malaysia (see also figures in Chan-ard et al. 1999; Grossmann 2006). A specimen from eastern Kalimantan showed a brown vertebral stripe and very small white dorsal flecks ( Grossmann 2006). Lim & Lim (1999) recorded G. s m i t h i i for the first time from Tioman Island, Malaysia (see Grossmann & Tillack 2004, 2005), which has been followed by Hien et al. (2001), and Grismer et al. (2002, 2004). However, the Pulau Tioman form differs from G. s m i t h i i sensu stricto according to Grossmann (2006) in eye coloration (turquous blue versus mossy green) and in ground coloration (gray brown versus green) and has been considered distinctive by Grismer (2006). In life, the form from Tioman more closely resembles G. v e r re a u x i, from which it differs in the lighter eye coloration (emerald green in G. verreauxi ) and the position of the nostril. We therefore suspect the form from Tioman Island to represent a distinct species and thus refer to it as Gekko sp. ”Tioman”. Additionally, Koch et al. (2009) published the first records of G. smihii from Sulawesi and Pulau Togian as well as from Pulau Batudaka. These authors argued against human introduction and rather believed that a natural dispersal from the Sunda shelf had occurred. Slight differences of the forms from Sulawesi and Togian compared with G. smithii s. str. were explained by Koch et al. (2009) as incipient divergence stemming from isolation, but they could not exclude the possibility that a cryptic species complex exists. Accordingly, we herein treat these both forms tentatively under the names Gekko sp. "Sulawesi" and Gekko sp. "Togian".
We herein consider the species of the Gekko gecko group a monophyletic unit. Species displaying green to blue or brick red iris (i.e., G. albofasciolatus , G. nutaphandi , G. siamensis , G. smithii , G. v e r re a u x i, Gekko sp. ”Tioman“, Gekko sp. ”Sulawesi“, Gekko sp. ”Togian“) and more-or-less green ground coloration seem to be closely allied. For remarks on Gekko gecko and congeners see below.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.