
Aoruroides chubudaigaku, Morffe & García & Hasegawa & Carreno, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4712.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:052D7A73-547D-4706-9BCC-98E141B4132F |
|
persistent identifier |
https://treatment.plazi.org/id/145A87F8-8E55-7656-FF51-FE9DFC88F853 |
|
treatment provided by |
Plazi |
|
scientific name |
Aoruroides chubudaigaku |
| status |
sp. nov. |
Aoruroides chubudaigaku n. sp.
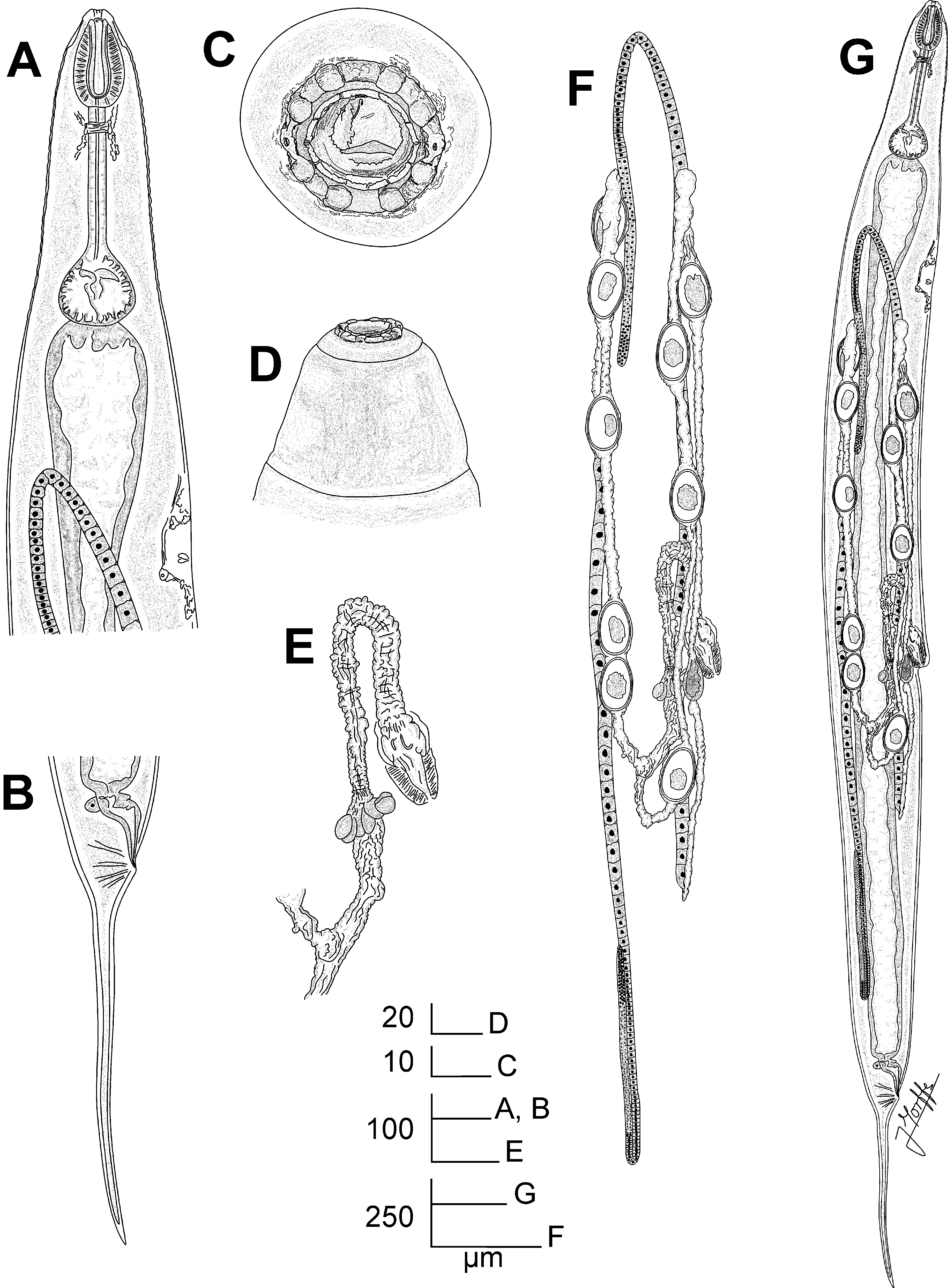
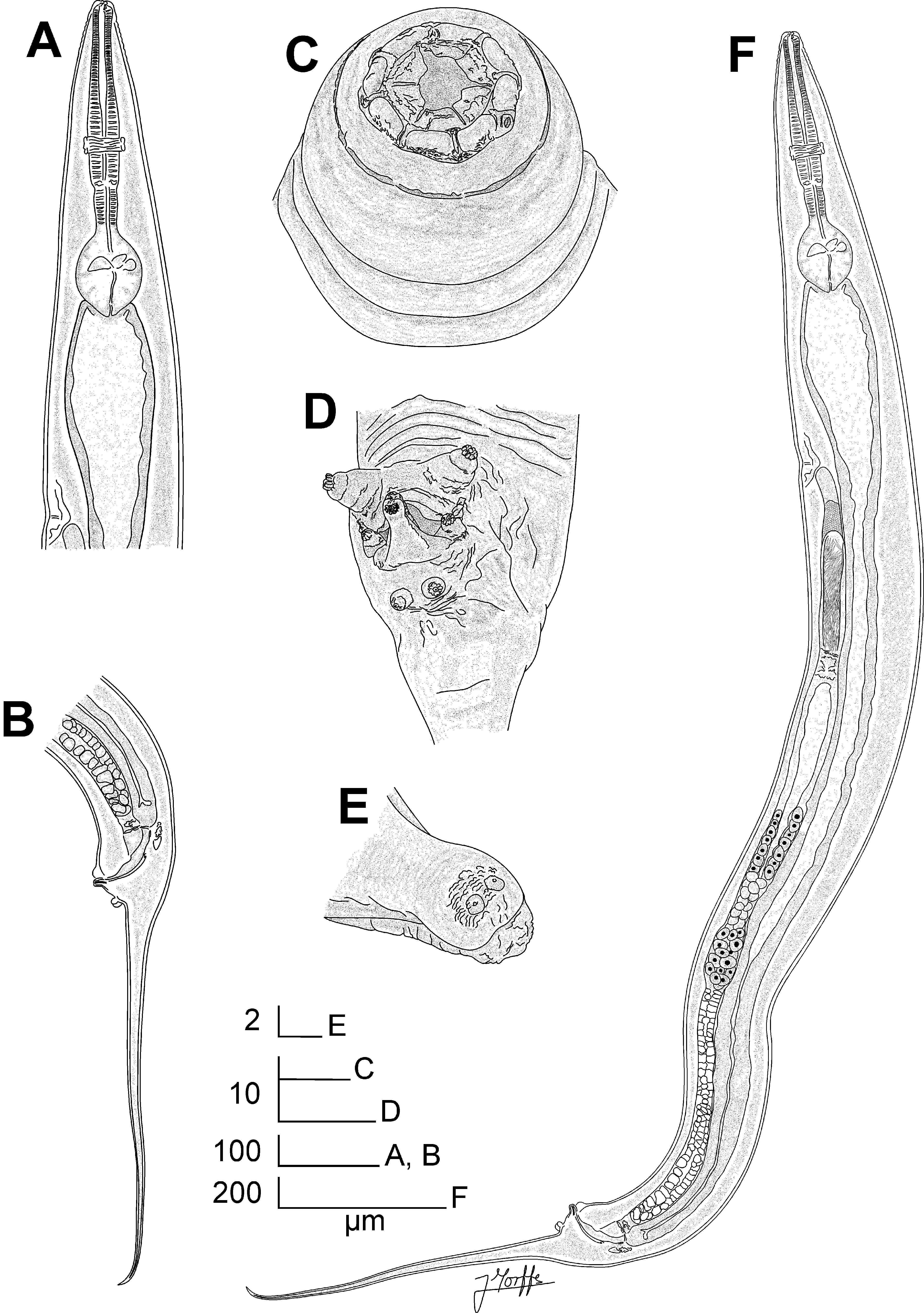
Fig. 1 View FIGURE 1 A-E, Fig. 2 View FIGURE 2 A-F, Fig. 3 View FIGURE 3 A-H
Type material. Holotype: ♀, Japan, Aichi prefecture, Kasugai, Chubu University Campus , 35º16´28.315´´N, 137º1´3.259´´E; in Panesthia angustipennis spadica ; X/2016; J. Morffe coll.; CZACC 11.7029 View Materials GoogleMaps . Paratypes: 5♀♀, same data as the latter; CZACC 11.7030 –11.7034 GoogleMaps . 5♂♂, same data as the latter; CZACC 11.7035 –11.7039 GoogleMaps . ♀, same data as the latter; NCHL-A17 GoogleMaps . ♂, same data as the latter; NCHL-A18 GoogleMaps .
Description. Female. Body robust, spindle-shaped, with the maximum body-width at level of the anterior lip of vulva. Cuticle with marked annuli in the cervical region, from the base of the first cephalic annule to midlevel of basal bulb, annuli ca. 14 µm. Lateral alae absent. Oral aperture lined by cuticular ridge, surrounded by eight rounded, paired cephalic papillae arranged as two sub-dorsal pairs and two sub-ventral pairs. The cephalic papillae are separated by elongated cuticular extensions. Amphids lateral in position, pore-like, closed by a valve-like structure. The amphids are located in cuticular prominences, connected to the cephalic papillae by slender cuticular extensions. First cephalic annule smooth, long, conoid and truncate, of ca. one third of the corpus length. Buccal capsule short and conoid. Oesophagus consists of a short, swollen, ovoid muscular corpus, notably set-off from the long (ca. 1.5 corpus-lengths long) cylindrical isthmus. Basal bulb rounded, valve-plate well developed. Intestine simple, sub-rectilinear, its anterior region slightly dilated. Rectum short. Anus with the posterior lip prominent. Nerve ring encircling the isthmus at the level of its first third. Excretory pore ventral, located at ca. one body-width posterior to the basal bulb. Vulva a ventro-median transverse slit, its anterior lip notably prominent, located near the level of the midbody. Vagina vera short, anteriorly directed. V agina uterina less muscular, extending anteriorly for ca. one vagina vera -length, reflexing posteriorly and continuing a short distance before joining to the uteri. One ring of rounded cells located at ca. the midpoint between the flexure of the vagina uterina and its link with the uteri. Genital tract didelphic-amphidelphic. Both ovaries reflexed. Anterior ovary reflexed at the level of the excretory pore. Posterior ovary reflexed to a distance of ca. one body-width before the level of the anus. A rounded spermatheca present in the posterior uterus, filled with spermatids. Oocytes in single rows. Eggs ellipsoidal, with the shell thin and smooth. Tail comparatively long, filiform, subulate, ending in a fine tip. Phasmids pore-like, lateral, located in the tail at ca. 20 µm posterior to the anus.
Male. Body smaller and less robust than females, its posterior end ventrally curved. Cervical cuticle unarmed. Lateral alae absent. Oral aperture surrounded by eight ellipsoidal cephalic papillae separated each other by short extensions of cuticle. Cephalic papillae not evidently paired and almost equidistant, except by the lateral cuticular extensions next to the amphids. Six labia projecting from the internal edge of the cephalic papillae through the oral aperture, arranged as two sub-dorsal, two sub-ventral and two lateral forms. Amphids pore-like, closed by a valvelike structure, lateral in position, located externally to the cephalic papillae in a cuticular prominence. First cephalic annule smooth, long, conoid and truncate, its length not surpassing the anterior dilation of the corpus. Oesophagus consists of a muscular, clavate corpus, set-off from the cylindrical isthmus, its anterior end slightly expanded. Basal bulb rounded, valve-plate well developed. Intestine simple, sub-rectilinear, its anterior region dilated. Nerve ring encircling the corpus at the level of its last third. Excretory pore ventral, located at ca. two body-widths posterior the basal bulb. Monorchic, testis reflexed at level of the excretory pore. Vas deferens divided into three regions: an anterior filled with rod-like spermatids, a median slightly swollen, its posterior part with rounded cells and a posterior region slender, with polygonal cells connected to the cloaca. Three pairs of copulatory papillae present: a pair of large, prominent, conoid pre-cloacal papillae in the anterior lip of cloaca; a pair of small adcloacal papillae located at both sides of the cloaca and a post-cloacal pair located at a short distance of the posterior lip of cloaca. Each papilla presenting eight small rounded projections surrounding the distal sensorial end. A single appendage located just posterior to cloaca, its base wide, becoming narrower toward the distal end (giving a triangle-like shape to the appendage) that is rounded and slightly dilated, with two minute papilla-like structures. Tail appendage comparatively long, filiform, subulate, ending in a sharp tip. Phasmids pore-like, lateral, located at level of the posteriormost pair of copulatory papillae.
Type locality. Chubu University Campus , Kasugai, Aichi prefecture, Japan, 35º16´28.315´´N, 137º1´3.259´´E GoogleMaps .
Type host. Panesthia angustipennis spadica (Shiraki, 1906) ( Blattodea : Blaberidae : Panesthiinae ).
Etymology. Specific epithet in apposition after Chubu Daigaku, Japanese for Chubu University. The campus of this university is the type locality of the species.
Differential diagnosis. Females of A. chubudaigaku n. sp. differ from A. cochinchinensis , A. legionarius Kloss, 1966 and A. queenslandensis by having the nerve ring at level of the first third of the isthmus vs. the midpoint of the isthmus. Also, A. cochinchinensis present a more anterior vulva than A. chubudaigaku n. sp. (V% = 46.50–47.30 vs. 47.02–51.93). A. legionarius can be differentiated from A. chubudaigaku n. sp. by its shorter body (2.664 –3.146 mm vs. 3.107 –4.361 mm), the comparatively shorter oesophagus (b = 8.33–9.45 vs. 6.99–8.62) and the comparatively longer tail (c = 2.88–2.90 vs. 4.43–6.64). The tail of A. chubudaigaku n. sp. is comparatively shorter than that of A. queenslandensis (c = 4.43–6.64 vs. 3.75–4.71).
The new species is similar to A. costaricensis Carreno & Tuhela, 2011 by the position of the nerve ring at the level of the first third of the isthmus ( Carreno & Tuhela 2011), the body length (3.107 –4.361 mm vs. 1.780 –4.070 mm) and comparative measurements of the oesophagus (b = 6.99–8.62 vs. 6.16–9.83) and tail (c = 4.43–6.64 vs. 2.91–4.52). They differ by the position of the excretory pore, located at a distance of ca. a body-width posterior to the basal bulb in A. chubudaigaku n. sp. vs. ca. half of the body-width in A. costaricensis . Also, the vulva is slightly anterior in A. costaricensis (V% = 46.18–46.19 vs. 47.02–51.93).
The females of A. rosario and A. travassosi Coy, García & Álvarez, 1993 are shorter than A. chubudaigaku n. sp. (1.620 –1.950 mm vs. 2.090 mm vs. 3.107 –4.361 mm) ( Coy et al. 1993a, b). The oesophagus (b = 3.91 vs. 6.99–8.62) and the tail (c = 3.37 vs. 4.43–6.64) are comparatively longer and the vulva is more anterior (V% = 46.89 vs. 47.02–51.93) in A. travassosi . The oesophagus of A. chubudaigaku n. sp. is slightly longer than that of A. rosario (b = 6.99–8.62 vs. 7.98–9.42). Aoruroides philippinensis tends to be longer than A. chubudaigaku n. sp. (4.000 –4.450 mm vs. 3.107 –4.361 mm) but its oesophagus is comparatively shorter (b = 8.24–8.46 vs. 6.99–8.62). Moreover, in A. philippinensis the vulva is slightly more anterior than in A. chubudaigaku n. sp. (V% = 45.50–48.99 vs. 47.02–51.93).
The males of A. chubudaigaku n. sp. can be easily differentiated from A. cochinchinensis and A. queenslandensis by lacking ornamentations in the cervical cuticle. Both aforementioned species present thorn-like and knob-like ornamentations, respectively. Moreover, the nerve ring is located in the posterior third of the corpus in A. chubudaigaku n. sp. instead of the union of the corpus with the isthmus in A. cochinchinensis and the midpoint of the corpus in A. queenslandensis . In spite of not being mentioned in the original description, the SEM pictures of the males of A. costaricensis show the presence of ornamentations in the cervical cuticle, in the form of minute scalelike projections in the annuli ( Carreno & Tuhela 2011). Such ornamentations are absent in A. chubudaigaku n. sp.
The nerve ring is located at the midpoint of the corpus in A. legionarius and A. philippinensis instead of the posterior third of the corpus as in A. chubudaigaku n. sp. Aoruroides legionarius has a longer body (2.386 –2.654 mm vs. 1.664 –2.108 mm) and the oesophagus is comparatively shorter (b = 7.80–8.29 vs. 5.62–6.53). Aoruroides philippinensis has a comparatively longer tail than A. chubudaigaku n. sp. (c = 1.86–3.88 vs. 3.67–5.37). The males of A. travassosi are the shortest among the genus (0.724 mm vs. 1.664 –2.108 mm) but their tail is comparatively longer than that of A. chubudaigaku n. sp. (c = 2.87 vs. 3.67–5.37).
DNA studies. Two partial sequences of the D2-D3 segment of the LSU rDNA were obtained from one female specimen (729 bp) and one male specimen (732 bp). Both sequences were identical, confirming the conspecificity of both sexes.
The other available sequences of species of the genus Aoruroides are three: one longer, belonging to A. cochinchinensis and comprising the D2-D3 domains of the LSU rDNA (as in the ones of the current study) and two shorter sequences of A. queenslandensis , only covering the D3 domain of the LSU rDNA ( Jex et al. 2006; Guzeeva et al. 2010). Aoruroides chubudaigaku n. sp. differs from A. cochinchinensis in 21 homologous positions in a 729 bp alignment. It differs from A. queenslandensis in five and six homologous positions for each sequence respectively, in a 274 bp alignment.
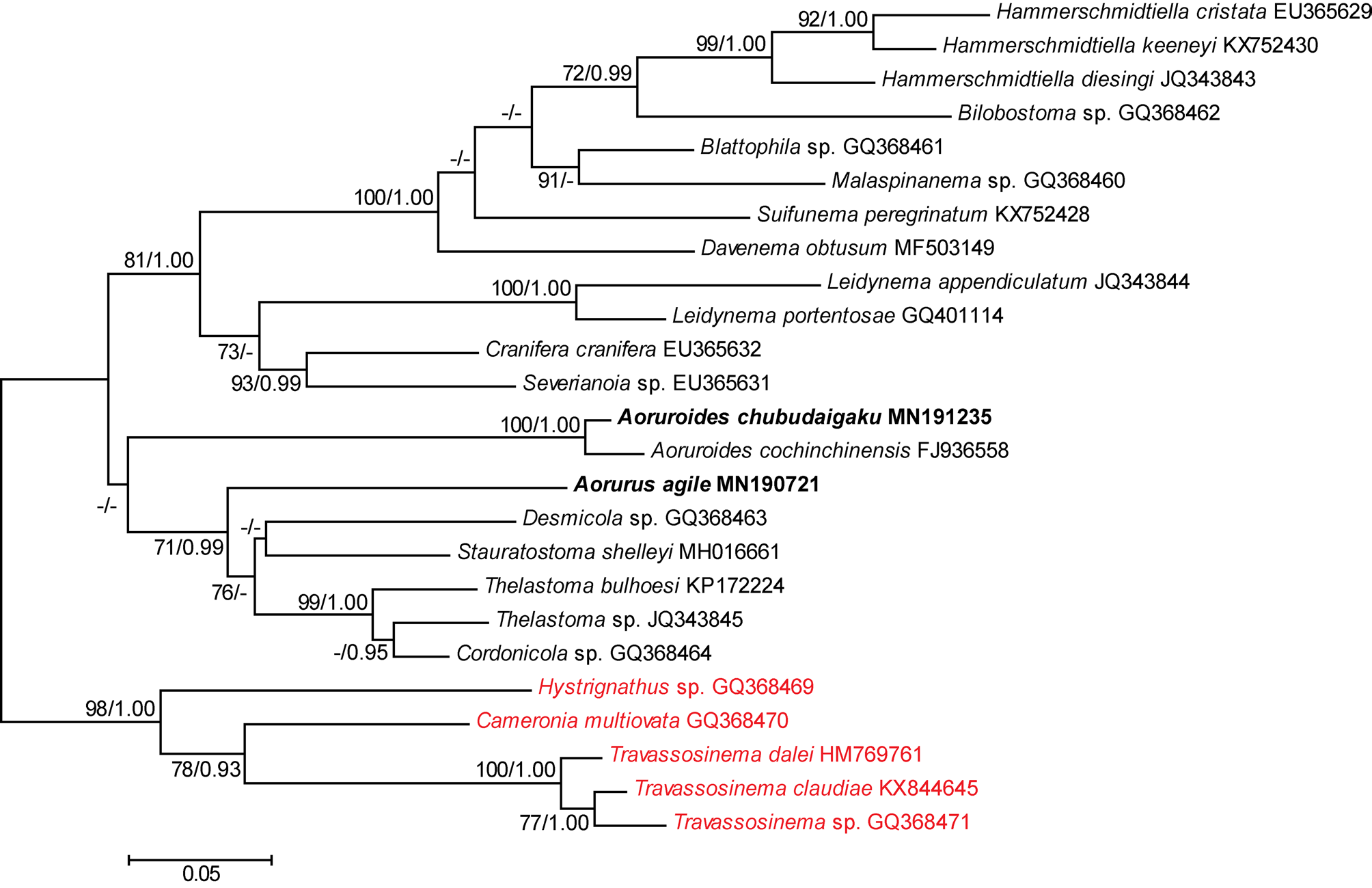
The topology of both, ML and BI trees was similar. Thus, only the ML is shown ( Fig. 4 View FIGURE 4 ). The two included species of Aoruroides form a monophyletic clade with high support values. This clade is located basally in a low-supported clade, formed by Thelastoma sp. + Cordonicola sp. + Thelastoma bulhoesi + Desmicola sp. + Stauratostoma shelleyi + A. agile . The latter species is also basally located in a monophyletic clade, formed by Thelastoma sp. + Cordonicola sp. + T. bulhoesi + Desmicola sp. + S. shelleyi .

| CZACC |
Coleccion Zoologia, Academia de Ciencias de Cuba |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Oxyuridomorpha |
|
Family |
|
|
Genus |