
Einfeldia australiensis (Freeman)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4158.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:6CC23B73-1DAC-4E48-BBE1-E035AA450E7B |
|
DOI |
https://doi.org/10.5281/zenodo.6060892 |
|
persistent identifier |
https://treatment.plazi.org/id/141487AE-AB25-FF9E-FF7B-69C8FB07FA5B |
|
treatment provided by |
Plazi |
|
scientific name |
Einfeldia australiensis (Freeman) |
| status |
|
Einfeldia australiensis (Freeman)
Chironomus (Xenochironomus) australiensis Freeman, 1961: 696 (adult male).
Type material. Holotype ♂, pinned, genitalia on celluloid, foreleg, head and antennae on separate slide in Euparal; AUSTRALIA: New South Wales, Oxford Falls , 3.iv.1922 (collector not stated) ( ANIC) . Paratypes (slide mounted in Euparal, Cranston): ♂, as holotype; ♂, N.S.W., Hornsby , 9.i.1958 (Lee).
Other material [all AUSTRALIA, and in ANIC unless stated otherwise]: Pe , Queensland, Fraser Island, Ocean Lake , 24°55.5’S 153°16.4’E, 2.vi.1997 (Wright, Reid, Gunn); 4L (3 slides) GoogleMaps , New South Wales, Sydney , Botany Wetlands, Pond #3, 33°56’08’’S 151°12’53’’E, 2.xi.2015, Sydney Water (SW) via Monica Mulder (2 slides, 1 larva on each; 1 slide, 2 larvae, with Procladius under 2 other coverslips (SW); 2L (one with chromosome squash) Blue Lagoon, 29o58’S 153 o15’E, 15.viii.1976 (Timms); 2Pe GoogleMaps , Australian Capital Territory, Jervis Bay , Lake McKenzie, 35°09’S 150°41’E, 28.iv.1997 (Wright); Pe, same except 7.iii.1998; Pe GoogleMaps , Victoria, Lake Little Beatle , 37°47.2’S 148°25’E, 17.xii.1996 (Wright) GoogleMaps ; Swan Lake , 38°12’S 141°19’E, 20.xii.1996 (Wright) GoogleMaps ; South Australia, Valley Lake , Mt. Gambier, 37o30’S 140o45’E, 15.viii.1972 (Timms), ‘ASA.34.2’; L (with chromosome squash) (Museum of Victoria) GoogleMaps ; 3L (with chromosome squash), Le / Pe / ♂, Pe / ♂, P(♂), Lake Edward via Kalangadoo, 37o38’S 140o35’E, 30.ix.1973 (Martin & Martin), ‘ASA 36.3’. GoogleMaps
Excluded from E. australiensis here (all = Xenochironomus spp.; see 'Remarks on Freeman's type series' below). Paratype, ♂, pinned, genitalia on celluloid, AUSTRALIA: [A.C.T.] Molongolo R.,. iii.1930 (Graham) . 2♂♂, 1♀ slide mounted in Euparal, several ♂, ♀, pinned, Northern Territory, Jim Jim Ck., WSW of Mt. Cahill , ‘at light’, 24.x.1972 (Colless).
Remaining four ♂ paratypes listed by Freeman (1961) not examined.
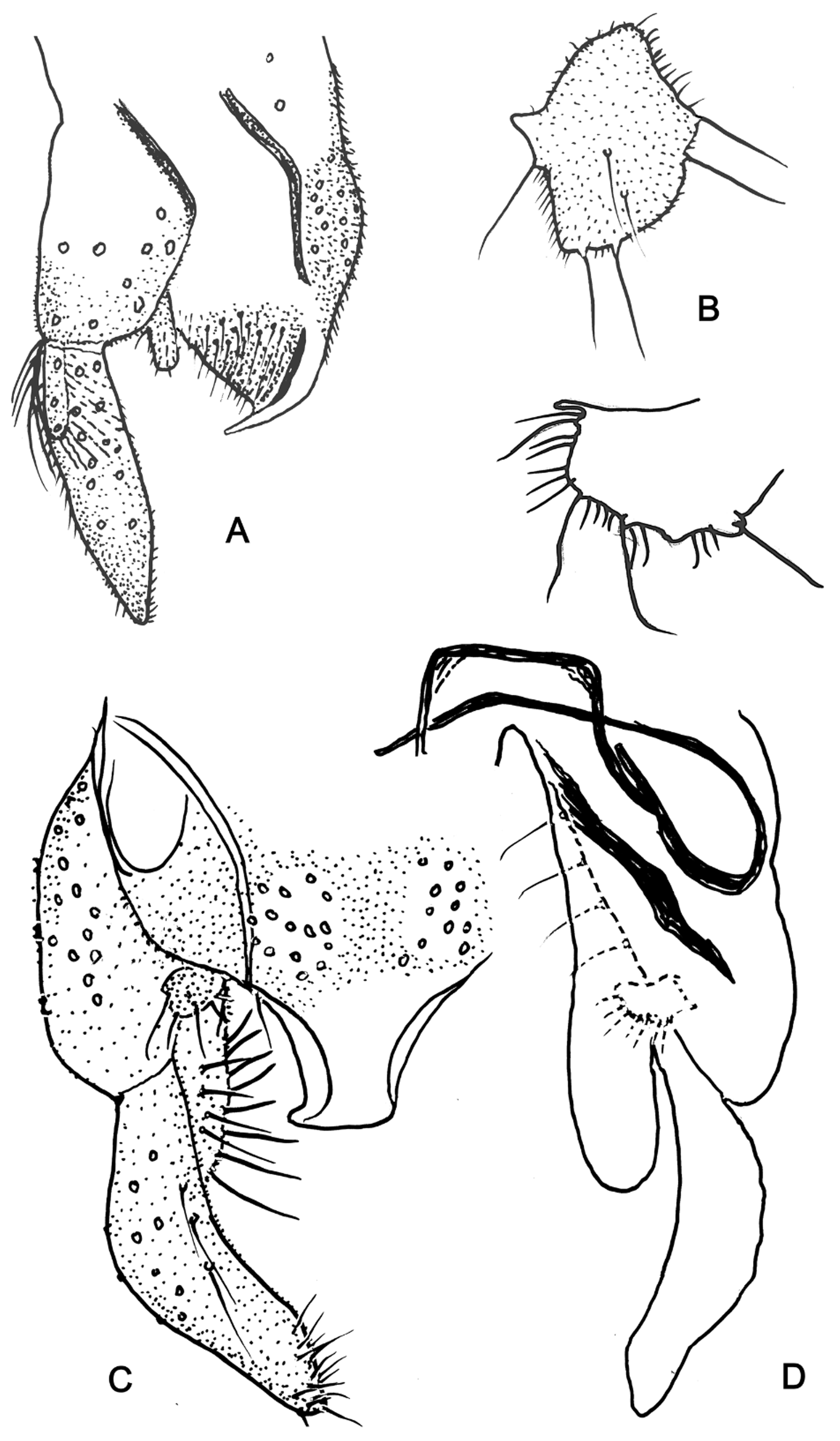
Description. Adult male ( Figs 1 View FIGURE 1 A, C, D; 2), (n=4, slide mounted, variously incomplete) [statements in square parentheses from Freeman (1961)].
Colour (dry, pinned) and size. Thorax yellow-brown with scarcely darker vittae; legs green, foreleg with darker tibia and tarsomeres, mid and hind legs with dark tarsomeres; abdomen light green becoming darker posteriorly [pale green, yellowish-red thoracic vittae, knees somewhat darkened, legs pale green, abdomen clear green]. Total length 5.0– 6.5 mm, wing length 2.2–2.9 mm (from arculus to tip) [2.75–3.5 (from origin at thorax to tip)].
Head. Antenna with 11 flagellomeres, fm 1–10 length 155–175 µm, fm 11 475–520; AR 2.7 [‘about 3’].
Frontal tubercle globular, 5–7 across [absent], without apical seta. Head with 15–26 irregularly biserial and continuous verticals + postorbitals; clypeus square, with 15–18 clypeals; palpomere lengths (2nd–5th) 50–65; 120– 175; 150–200; 210–320; 4–6 linearly arranged sensilla chaetica on 3rd segment, not arising from pit.
Thorax. Antepronotal lobes quite narrow dorsally and fused at shallow notch ( Fig. 1 View FIGURE 1 A), without antepronotals; mesonotum with slight indication of scutal tubercle; acrostichals 19–21 in irregularly biserial row starting near antepronotum, 6–11 uniserial dorsocentrals, 5–6 long prealars, c. 16 biserial scutellars of 2 sizes.
Legs variously damaged: Leg Ratios: foreleg 1.57–1.65 [nearly 2], mid leg 0.46–0.48, hind leg 0.55–0.6. Foreleg BV ratio 0.63, SV ratio 0.65 (both from holotype only). Fore tarsal beard sparse, BR 5.0 on tarsomeres 1 and 2. Fore tibial scale low, rounded without spur ( Fig. 1 View FIGURE 1 C); each mid and hind tibial apex with wide and narrower comb, each comb with 25 long spur ( Fig. 1 View FIGURE 1 D). Tarsal sensilla chaetica (n=1) 26 linear in distal 2/3 of ta1 of mid leg, 13 in distal ¼ of ta1 of hind leg. Fore tarsomere 5 cylindrical. Pulvilli strong, subequal to claw.
Wing generally as in Cranston et al. (1989: fig. 10.19B), plain, membrane with only fine punctation; R2+3 running close to R1, ending close to apex of R1, substantially prior to wing apex; R4+5 and C ending together near wing apex, without costal extension. Wing vein setation: R with 25–33, R1 with 17–26 along complete length, R4+5 with 20–34 in distal half; 8–16 squamals. VR (ratio cubitus to media) 1.07.
Genitalia ( Fig. 2 View FIGURE 2 ). Laterosternite IX without setae. Tergite IX with anal tergite bands moderately developed, widely separated and ending distally near origin of anal point. Fig. 2 View FIGURE 2 A, C); dorsomedial TIX setae in pair of clusters between the tergite bands, 11–20 setae in each cluster. Anal point ( Fig. 2 View FIGURE 2 A, C) arising anteriorly of posteroventral end of tergite IX, very broad and variably curved ventrad ( Fig. 2 View FIGURE 2 A) such that its length can be impossible to measure in dorsal view; apparently T-shaped in cross-section; dorsal surface bare. Superior volsella ( Fig. 2 View FIGURE 2 A–C) in dorsal view trapezoidal or pad-like, entirely trichiose (not shown in Fig. 2 View FIGURE 2 B, bottom), with variably located shorter to longer setae on dorsal, medial, caudal and lateral faces; volsella laterally with variably developed short, subacute process. Inferior volsella ( Fig. 2 View FIGURE 2 C, D) elongate, parallel-sided, not clubbed distally, microtrichiose and with many distal setae, but without any special terminal seta. Gonocoxite 185–210 long, with medial row of 4– 5 setae ( Fig. 2 View FIGURE 2 D). Gonostylus 175–220 long, slightly widened in mid-section, its tip tapering slightly, medially bearing 8–10 stronger, clustered setae ( Fig. 2 View FIGURE 2 C). Major internal apodemes of segment IX as in Figure 2 View FIGURE 2 D.
Female unknown.
Pupa (Fig. 3) (n=8). Length c. 7 mm, medium brown, with weak abdominal apophyses.
Cephalothorax. Frontal warts absent. Cephalic tubercle (Fig. 3A) broadly conical, c. 75 long, with short (20 µm), pale frontal seta inserted slightly subterminally. Thorax rugose especially anterodorsally, with weak scutal tubercle. Thoracic horn multi-branched, hyaline, arising from double, medially narrowly divided tracheal plate in medially slightly constricted basal ring (Fig. 3B). Prealar area with two projecting lobes, anterior lobe directed laterally, posterior one more posteriorly (Fig. 3C).
Abdomen. Tergal armament (Fig. 3D): Tergite I with posterior transverse fine shagreen; TII–VI more broadly and densely spinulose in near continuous central patch, including pair of posteromedial patches with larger points (Fig. 3D); spinulose area larger on more posterior segments; TVII with anterolateral areas of fine spinules, TVIII– IX bare. Hook row on TII undivided, with 55–60 uniserial hooks, extending c. 50% width of segment. Paratergites II–IV spinulose, others bare. Conjunctives all bare. Sternite I with no lateral or medial anterolateral tubercles, with sparse fine posterior spinulation. SII with posteromedial pattern of short to elongate and strong but translucent spines (Fig. 3E). Remaining sternites bare. Posterolateral 'comb' of segment VIII (Fig. 3G) of 5–7 brown, basally separated, tapering spines c. 80–110 long. Anal lobe with dense, uniserial fringe of 70–80 taeniae, with single dorsal seta inserted quite anterolaterally. Pedes spurii A on SIV strongly developed. Pedes spurii B well-developed on II, absent on III.
Setation: 1 pair of anterolateral O setae each on tergites II–VIII and apparently on at least anterior sternites. Tergite I with 2 D setae, II with 4, III–VII with 5, VIII with 1. Sternite I with 2 V setae, II–VII with 3–4, VIII with 1. L setae on I–IV semi-taeniate, 1 on I, 3 on II with the L3 located ventrally on bases of pedes spurii B; 3 each on III and IV, the L3 exceptionally on respective conjunctive. 4 taeniate L setae each on V–VIII.
Larva ( Fig. 4 View FIGURE 4 ). Length c. 7–12.5 mm, head capsule length c. 660–700, postmentum length 270–280. Head pale yellow, with occipital margin, teeth of mentum, ventral teeth of mandible and premandibular teeth variably contrasting brown. Eyespots double, dorsal spot above or slightly anterior of ventral one, both more or less circular. Dorsal surface of head ( Fig. 4 View FIGURE 4 A) including a fused frontoclypeus with cephalic setae S5 inserted anteromedially of dorso-lateral pits, with large, ovoid to heart-shaped fenestra between the S4, with S3 inserted subapically in broad ‘scalloped’ area representing fused clypeus, with convex anterior margin. Lateral sclerites granulose, fragmented and not individually distinguishable; S2 arising in that area near posterolateral corners of labral sclerite. Labral sclerite strongly widened anteriorly, with S1 arising near the mid-lateral bend.
Antenna ( Fig. 4 View FIGURE 4 B) 5-segmented with segment lengths 88–93: 25–28: 12–15: 12–15: 7; AR 1.4–1.6; Lauterborn organs slender, opposite, 10 long; style slender, 10 long; blade 65–70 long, extending to antennal apex. Labroepipharyngeal region ( Fig. 4 View FIGURE 4 C, D): SI setae finely plumose along most of both edges, SII simple. Pecten epipharyngis comprising three indistinctly separated scales, each scale with 4–8 teeth of somewhat variable shape, size and level of origination on plate ( Fig. 4 View FIGURE 4 D), premandible 100–105 long, with 2 long apical teeth and two short, partially fused proximal teeth; brush extensive. Mandible ( Fig. 4 View FIGURE 4 E, F) 215–225 long, with short, pale brown dorsal tooth and small (15 long), more proximal accessory dorsal tooth ( Fig. 4 View FIGURE 4 F); apical tooth and three inner teeth dark, innermost tooth clearly delimited from mola; seta subdentalis 15 long, hooked apically; mola smooth, without spinules on medial edge. Maxilla ( Fig. 4 View FIGURE 4 G) with chaetulae of palpiger c. 15 long.
Mentum ( Fig. 4 View FIGURE 4 H, I) 155–160 wide; single median tooth domed, with pair of variably visible lateral accessory teeth (strongly worn in Fig. 4 View FIGURE 4 I); each side of mentum with six lateral teeth evenly decreasing in size. Ventromental plate ( Fig. 4 View FIGURE 4 J) 165–175 wide, 80 deep, with 40–46 striae extending near full depth and across complete width of plate, each stria terminating anteriorly in 1–2 spinules slightly posterior to plate margin.
Abdomen. Anterior parapod with dense cluster of pale golden, simple claws. Abdominal segment VIII with one pair of curved or partially coiled ventral tubules c. 1.1–1.6 mm long. Procerci and posterodorsal ('supraanal') setae of segment IX pale to medium brown, procercus with 6–7 brown, c. 800 long anal setae. Posterior parapods 390–420 long, claws golden. Anal tubules pale, sausage-shaped without constriction, c. 2x as long (250–320) as wide.
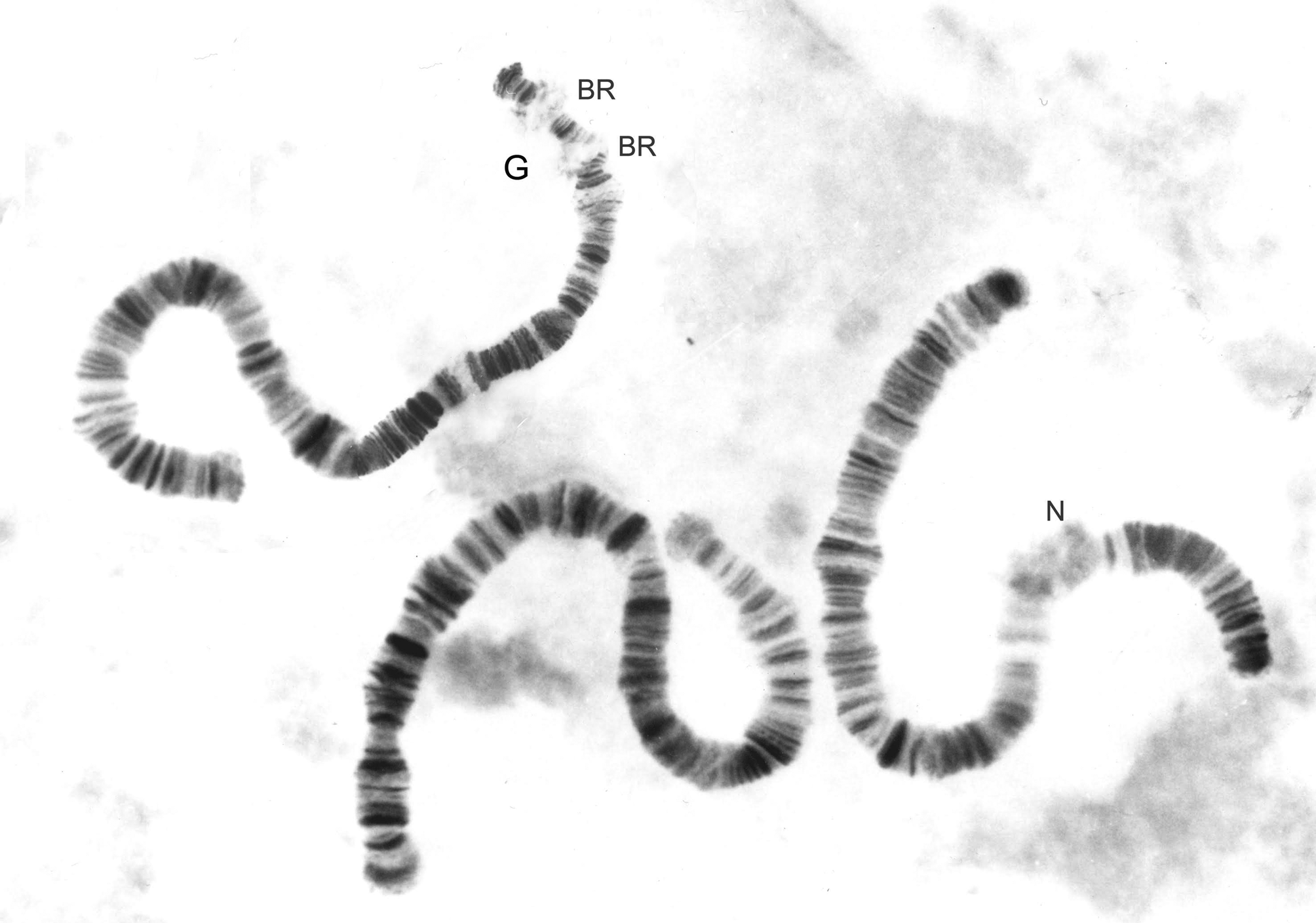
Cytology. The karyotype comprises three polytene chromosomes ( Fig. 5 View FIGURE 5 ). Comparison to the karyotypes of two North American Einfeldia species ( Martin 2016) indicates that this is derived from a typical 2n=4 karyotype, comprising seven chromosome arms, common to most species of Chironomus (s. l.) ( Michailova 1989, Martin 1996), i.e. the small acrocentric chromosome is fused to the end of one arm of a metacentric chromosome. The banding pattern cannot be compared directly with those of Chironomus (Keyl 1962, Devai et al. 1989), although there are some hints of general homology for some chromosome arms. Almost certainly the terminal segment bearing two Balbiani rings (BRs) is the equivalent of arm G and this may be fused to the equivalent of arm E. The position of the centromere is unclear in this chromosome, whereas in the other two chromosomes there are heterochromatic bands that are likely to represent the centromeres. The nucleolus is near the middle of the arm of one of these two chromosomes. No polymorphisms were present in the studied individuals.
Ecology and significance to biomonitoring. Enough records of the immature stages of E. australiensis have been found to allow a summary of the distribution and assess the types of water bodies in which they live. Pupal records dominate—the stage is distinctive, easily collected and has been recognised in lentic studies (e.g. Wright & Cranston 2000, Wright & Burgin 2007). In South Australia, Lake Edward is a productive maar lake with surface area 29 ha, maximum depth 7 m and mean depth 4.2 m, probably unstratified, with black mud sediment comprising 46.6% organic matter, 30% carbon and 1.8% nitrogen ( Timms 1974). In this lake, larvae of ‘ X. ’ australiensis were found in one of eight samplings amongst weed at a depth of 1.5 m (leg. et det. J. Martin, August 1973). The same collector reported larvae also from nearby Valley Lake (37"51’S; 140º46' E) at a depth of 10 m and a density of 17 individuals m -1.
In subcoastal northern New South Wales, larvae of ‘ X.? australiensis ’ were reported by Timms (1982) from Blue Lake, a freshwater lake with marine contact and variable water chemistry according to season and status of the marine connection. Further south in Jervis Bay, Lake McKenzie is a permanent, closed, variably shallow (max. 9 m deep) freshwater dune lake of 9 ha, with pH 5.5–6.5, relatively dilute (160–230 µS/cm), with P 30 pg/l and N below detection limits, and with thermal stratification when sampled. Although ‘ X. australiensis’ was not reported from there by Norris et al. (1993), exuviae were found by Wright & Cranston (2000) to be common in late austral summer (April) but absent in cooler September.
Recently larvae were collected from Botany Wetlands, eastern Sydney, where sediment conditions are essentially sand with a thin layer of biological matter. The water was slightly turbid at the time of collection but usually it is quite clear for a ‘wetland’. Conductivities in the connected Pond 5 and Mill Pond typically range between 100 and 400 µS -1 but reached 530 µS.cm -1 on occasion, indicating an intermittent marine connection. High conductivities, suggesting marine incursions, are noted also at Ocean Lake, a disturbed dune lake on Fraser Island, Queensland, where conductivity was 365 µS.cm -1 and pH 6.3 (Ian Wright, pers. comm. 2016).
In summary, the immature stages of E. australiensis occur in relatively shallow mesotrophic to eutrophic, circum-neutral pH and high-conductivity dune lakes and maars, from southeastern Queensland to southern Australia.
Remarks on Freeman's type series. The type series of C. (X.) australiensis comprises pinned adults, several of them with a genitalia preparation on a celluloid mount associated with the pinned specimen (i.e. on the same pin). The holotype is located in ANIC, as are several paratypes, including from the type locality, Oxford Falls, NSW. Freeman (1961) stated the holotype and 3 paratypes to be in ‘SPHTM’ (the School of Public Health and Tropical Medicine, University of Sydney), but this collection has been de-accessioned and divided between other institutions, including ANIC. Some other paratypes were listed as in CSIRO, but not all of these are present there.
The extant material is mixed: the holotype and some paratypes belong to Einfeldia , but at least one paratype (from Molongolo R.) belongs to an undescribed species of Xenochironomus . A series of adults from Northern Territory, identified subsequently by D.H. Colless, all belong to Xenochironomus . The respective adults resemble each other, differing subtly as elaborated below, but can be separated as follows. The antepronotal lobes are narrowly fused medially in the Einfeldia specimens, whereas they make slight contact but are weakly separated in the Xenochironomus ; frontal tubercles are present in the Einfeldia , absent in the Xenochironomus ; the superior volsella is subrectangular with a small posterolateral projection in the Einfeldia , but tongue-shaped and without projection in Xenochironomus .
Transferring C. (X.) australiensis to Einfeldia does not remove the modestly diverse but rarely-reared genus Xenochironomus from Australia. Aside from the paratype and N.T. specimens discussed above, we are aware of adults of at least two unnamed species, several forms of pupal exuviae and larvae ( Cranston 1996).


| ANIC |
Australian National Insect Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Einfeldia australiensis (Freeman)
| Cranston, Peter S., Martin, Jon, Mulder, Monica & Spies, Martin 2016 |
Chironomus (Xenochironomus) australiensis
| Freeman 1961: 696 |