
Eulamprotes wilkella (Linnaeus, 1758)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3746.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:A34057EB-13C1-4E5D-BF39-0AD1EE10AB12 |
|
DOI |
https://doi.org/10.5281/zenodo.6146577 |
|
persistent identifier |
https://treatment.plazi.org/id/13174807-7A1D-FFF3-E98C-F8FD1F10E0EF |
|
treatment provided by |
Plazi |
|
scientific name |
Eulamprotes wilkella (Linnaeus, 1758) |
| status |
|
Eulamprotes wilkella (Linnaeus, 1758)
Phalaena (Tinea) wilkella Linnaeus 1758: 541 . Phalaena (Tinea) merianella Linnaeus 1758: 541 . Tinea germarella Geyer 1832 : pl. 70.
Gelechia (Brachmia) pictella Zeller 1839: 202 . Gelechia tarquiniella Stainton, 1862: 112 .
Examined material. Andorra (n=1), Austria (n=17), Bulgaria (n=2); Denmark (n=265), Finland (n=3), France (n=19), Germany (n=3), Great Britan ( England) (n=2), Greece (n=5), Hungary (n=1), Italy (n=43), Macedonia (n=1), Netherlands (n=3); Norway (n=3), Poland (n=2), Romania (n=1), Spain (n=9), Sweden (n=48), Switzerland (n=2).
Kazakhstan: 2♂, Zailiskiy, Alatau, Alma-Atinskij Nat. Par., 43˚05’N, 77˚15E, 1650 m, 14.vii.1990, leg. Kaila & Mikkola (ZMUH); 2♂, Dzhambulskaya obl., 70 km NNE Frunze, 43˚24’N, 75˚02E, 950 m, 18.vii.1990, leg. Kaila & Mikkola (ZMUH);
Kyrgizia: 5♂, 30 km E Naryn, 41˚25’N, 76˚20E, 2500 m, 22.–29.vii.1990, leg. Kaila & Mikkola (ZMUH); 1♂, 41 km E Naryn, 41˚26’N, 76˚29E, 2500 m, 9.viii.1990, leg. Kaila & Mikkola (ZMUH); 5♂, 40 km W Lake Issyk Kul, leg. Kaila, 13.viii.1990, leg. Kaila & Mikkola (ZMUH);
Russia: 3♂, Irkutsk oblast, 20 km S Ust-Ordynski, leg. Sinev (ZIP); 1♂, Tuva Republic, E. Tannu-Ola Mts., 5 km ENE Khol-Oozha, 50˚45’N, 94 ˚29’E, 2175 m, 17.vi.1995, leg. Jalava & Kullberg (ZMUH); 2♂, Irkutsk obl., 40 km N Irkutsk, 1.–3.viii.1983, leg. Mikkola & Vitiasaari (ZMUH); 1♂, Sakha Republic, 147 km SW Jakutsk, Elanka, 8.–11.vii.1979, leg. Ammosov (ZMUH); 1♂, Southern Primorje, Andreevka, 10.–15.viii.1998, leg. Jalava & Kullberg (ZMUH).
Turkey: 3♂, Ankara, 10 km N Kizilcahaman, 1150–1250 m, 6.–7.viii.1989, leg. Fibiger (ZMUC).
DESCRIPTION.—Adult ( Figs 1–3 View FIGURES 1 – 12 ). Male. Wingspan 10–12 mm. Labial palpus cream-white; basal third of segment 2 blackish brown on outer surface; tip of segment 3 black. Basal part of antenna blackish, becoming more distinctly lighter ringed in middle; apical quarter white. Head yellowish white with lighter frons and a dark line around eye; thorax and tegula black. Forewing black with three silvery white fasciae: first oblique from 1/6 at costa to 1/5 at fold; second from middle of costa reaching two-thirds towards dorsum; an inwards directed fascia from a cream-white pre-apical costal spot to tornal spot, often slightly interrupted; some silvery white scales in apex and along termen; cilia blackish grey, whitish grey at tip of apex. Hindwing almost as broad as forewing, light grey with grey cilia. Abdomen yellowish grey in basal third, then dark grey with yellow-white tip.
Female. Wingspan 8–11 mm. Hindwing about two-thirds as broad as forewing. Otherwise similar to male.
Variation. The fascia from pre-apical costal spot to tornal spot can be straight or slightly angled, and it can be interrupted or unbroken. There is some variation in the distinctness of light and dark rings in the basal threequarters of the antenna, but the apical quarter is always white. Specimens from some British and Irish localities have the silvery white markings in the forewings reduced to only one (f. tarquiniella ) ( Fig. 2 View FIGURES 1 – 12 ) or two fasciae. There seems to be also some variation in the wing form of females, with a tendency to (slight) reduction in breadth of hindwing and more or less acuminate forewings (see also Heckford 2002).
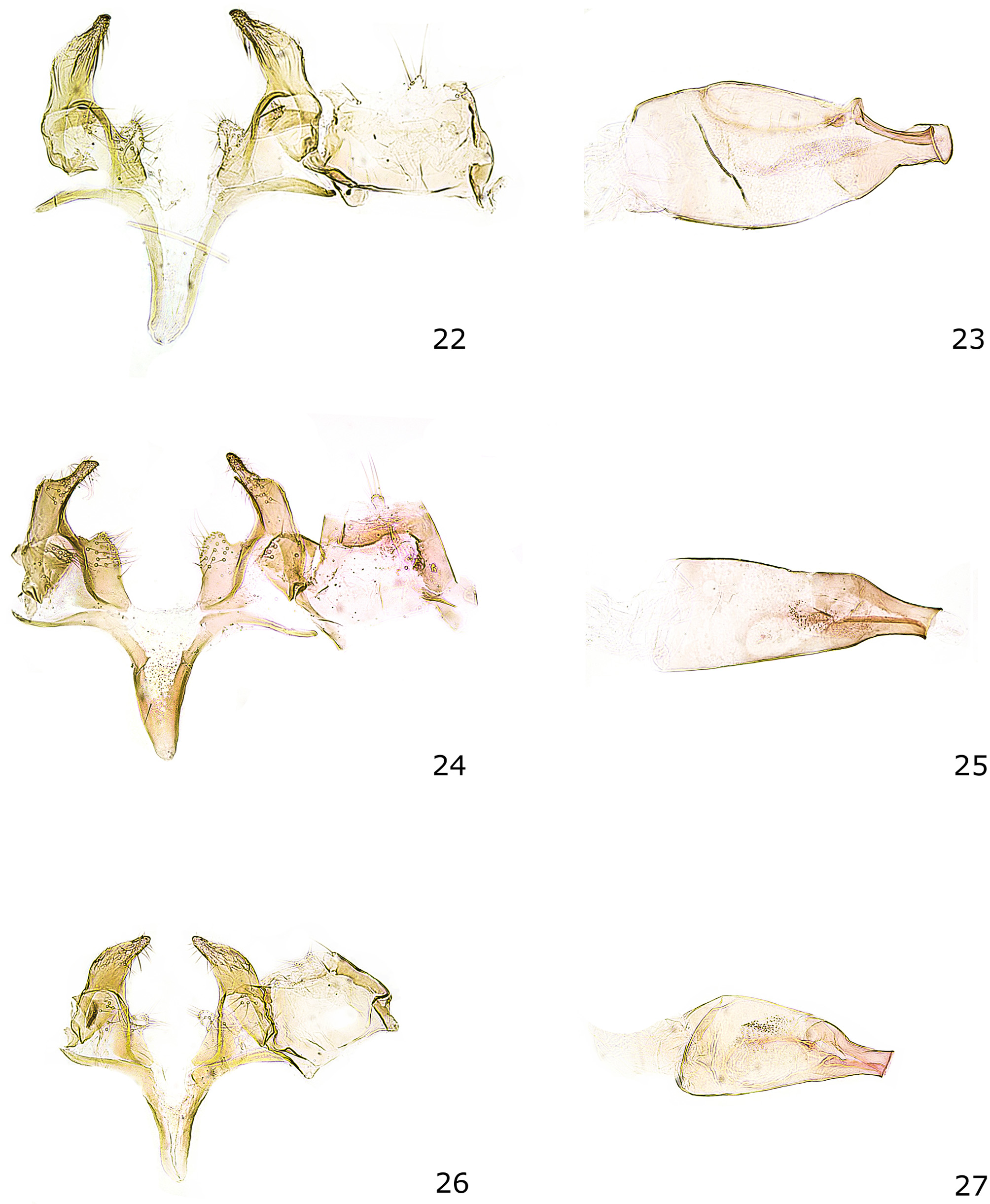
Male genitalia ( Figs 22–23 View FIGURES 22 – 27 ).—Segment VIII with two pairs of coremata in intersegmental membrane, grouped into short tufts of moderately broad and lanceolate scales, respectively. Uncus with tiny process, apically with three long and few short setae; tegumen short, sub-rectangular, a sclerotized belt (gnathos) connects the dorsolateral corners of the tegumen, from these corners two very long and rather stout setae are arising, anterior margin nearly straight, pedunculi small; valva has shape of bird's head in lateral projection, moderately slender, with strongly convex mediodorsal (outer) and nearly straight ventral (inner) margin, distal part strongly tapered to pointed apex; separate plate-like sclerite at base of valva, covered with few setae; sacculus a broad, setose lobe; saccus about as long as distance from anterior margin of vinculum to tip of valva, basally moderately broad, distally evenly tapered; phallus broadly bulbous, apical fifth abruptly and strongly tapered, about one quarter of maximum width of its anterior part; vesica with number of small grains.
Female genitalia ( Fig. 50 View FIGURES 50 – 51 ).—Apophysis posterioris slender, rod-shaped, about three times length of segment VIII; segment VIII with distinctly sclerotized posteriolateral part, otherwise membranous; ostium bursae laterally with sclerotized folds; apophysis anterioris slender, rod-shaped, about 2.5 times as long as segment VIII; ductus bursae long and slender, posterior half with slender, oblong sclerite extending distinctly beyond apophysis anterioris, anterior half membranous, weakly expanded; corpus bursae sub-oval, slightly larger than segment VIII; signum a large, nearly sickle-shaped transverse plate, anterior margin with long, posterior edge with shorter stiletto-shaped process.
DIAGNOSIS.— E. wilkella is characterized by having the apical quarter of the antennae white, and the pre-apical costal spot and the tornal spot of the forewing are (normally) fused. For E. wieseri sp. nov., E. altaicella sp. nov. and E. libertinella see under these species. The almost completely reduced uncus, the slender distal part of the valva and the strongly inflated phallus are collectively diagnostic, though particularly the differences from E.
superbella are small with a less stout valva in E. wilkella and a differently shaped phallus. The female genitalia differ from all other known species in the nearly sickle-shaped of the signum.
GENETIC DATA.—The intraspecific divergence of the barcode region ranges from 0%–1.38% (average distance 0.62%) (n=10). The distance to the nearest neighbour E. superbella is 7.48%.
DISTRIBUTION.—Almost all of Europe; Turkey; Russia: Siberia. Records from China (Li 2002) and Korea (Park & Ponomarenko 2007) need verification. For records from Japan (Sakamaki 1995) see below under remarks. The form with reduced markings in the forewing is only known from a few localities in Great Britain and Ireland.
BIOLOGY.—The larva is described by Stainton (1867) and Sterling (2008). It has been reported feeding on Cerastium fontanum Baumg. (Stainton 1867) , an observation which was widely accepted and adopted even in recent standard literature (i.e. Elsner et al. 1999). However, Sterling (2008) found that the larva feeds on the leaves of the moss Syntrichia ruraliformis (Besch.) Cardot , living in silken tubes at the base of a stem of the foodplant, extending into the sandy soil/moss mix beneath. Sterling (op cit.) suggests that Cerastium L. is only an exceptional host for E. wilkella ; this may also have been the case with one specimen which emerged from Cerastium plants collected early May at the island Møn, Denmark, together with several specimens of Caryocolum alsinella (Zeller) (O. Karsholt, unpublished). According to Sterling (op cit.) the larval period lasts from March to May and a second generation as stated by Stainton (1867) could not be verified. Continental authors such as Elsner et al. (1999) suggested bivoltinism, with moths flying from May to June and again from July to September but voltinism is unconfirmed and the flight period may refer to one long lasting generation only. The species seems to prefer dry habitats, coastal as well as inland, but it is also found in other types of localities like woods, gardens and parks. Males are easily attracted to light, whereas females are mostly found by sweeping vegetation during day, often where Thymus is flowering.
REMARKS.— Phalaena (Tinea) wilkella was described from an unspecified number of specimens. Robinson & Nielsen (1983) designated a male from Gotland ( Sweden) as the lectotype and synonymized Phalaena (Tinea) merianella Linnaeus, 1758 , described on the same page immediately before, with P. wilkella . Even though this interpretation of Ph. merianella can be disputed as Linnaeus’ description differs from that of Ph. wilkella it serves stability best to follow the synonymization by Robinson & Nielsen (op cit.).
The identity of Tinea germarella has been disputed and we follow here the prevailing usage as a junior synonym of E. wilkella (Huemer & Karsholt 2011) .
Gelechia (Brachmia) pictella was described from three specimens collected in August on sandy soil near Glogow ( Poland) (Zeller 1839) and synonymised with E. wilkella by Sattler (1973).
Gelechia tarquiniella was described from an unspecified number of specimens of both sexes collected in June in the sandhills near Dublin ( Ireland) (Barrett 1861, Stainton 1862) and later synonymized with E. wilkella by Meyrick (1928). From the original description it is clear that the name tarquiniella refers to the form with only a basal posterior bar. The form where only the posterior fasciae of the forewings are missing is unnamed. Bond (1991) came to the opposite conclusion, but the specimen referred to by him as “Stainton’s type specimen” is apparently not part of the type series (K. Sattler in litt.).
Sakamaki (1995) recorded E. wilkella as new to Japan, based on comparison with specimens from North Europe. In his description of Japanese specimens he did not mention the characteristic white tips of the antennae and, according to his description, the labial palpi differ from those of E. wilkella in having segment 2 (and the head) yellow, whereas segment 3 is black. We examined one similar, but rather worn male from Japan, and three such specimens from Far East Russia (in ZIAP and ZMUH). We find it likely that they belong to a further species related to E. wilkella , but due to the condition of the examined material we refrain from describing it here.
We examined at least one correctly identified specimen of E. wilkella from Far East Russia (see above).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |