
Dioscorea mayottensis Wilkin, 2007
|
publication ID |
https://doi.org/10.5281/zenodo.5187427 |
|
persistent identifier |
https://treatment.plazi.org/id/11354F5E-FFDA-FF9D-7A8D-FA2EA4516E88 |
|
treatment provided by |
Carolina |
|
scientific name |
Dioscorea mayottensis Wilkin |
| status |
sp. nov. |
Dioscorea mayottensis Wilkin View in CoL , sp. nov.
( Figs 2-4 View FIG View FIG View FIG )
Dioscorea comorensi affinis sed pubescentia fugaci atque in gemmis juvenilibus tantum inventa (nec magis persistenti in surculis juvenilibus atque axillis inventa), inflorescentia mascula e cymulis alternis ordinatim dispositis 4.5-14 mm distantibus constanti (nec cymulis irregulariter dispositis nec suboppositis 0.7-5.0[-7.3] mm distantibus), ramis cymulae pedicellisque tenuioribus, pedicello in floris primi cymulae 6 mm vel plus longo, pedicello florum sequentium 4.3 mm vel plus longo (nec 3.1 mm longo vel minus), tepalis omnibus similibus (nec interiore latiore quam exteriore), filamentis spiralibus vel contortis (nec reflexibus) differt.
TYPUS. — Mayotte. Grande Terre, Bandrélé, ♀ fl. & young fr., 9.II.2001, Barthelat, M’Changama & Sifary 296 (holo-, P!; iso-, G, K!, MAYOTTE, MO) .
PARATYPES. — Mayotte. Grande Terre , Saziley , Réserve forestière de Saziley, ♀ fl., 23.I.2001, Barthelat , M’Changama & Sifary 258 (G, K!, MAYOTTE, MO, P!). — Sazilé Bé , pied de falaise, 12°58’39”S, 45°12’01”E, ♀ fl. & young fr., 11.IV.1999, Pignal, Pibot & Soumille 1125 (K!, MO, P!). — Sazilé Bé, pied de falaise, 12°58’39”S, 45°12’01”E, ♂ fl., 11.IV.1999, Pignal, Pibot & Soumille 1126 (B, NY, P!). — Pointe Handrema, ♀ fr., 22.IV.1999, Mas 40 (P!). — Pointe Handrema, ♂ fl., 22.IV.1999, Mas 64 (P!). — Îlot de Baudilé [Îlot de Bandrélé], ♀ young fr., 30.III.1975, Coulon 28 (P!). — Îlot de Baudilé [Îlot de Bandrélé], ♂ fl., 30.III.1975, Coulon 31 (P!). — Grande Terre, Combani, Mont Combani, Réserve forestière, ♀ fl. and young fr., 10.I.2002, Barthelat, M’Changama & Ali Sifari 670 (G, K!, MAYOTTE, MO, P!). — Grande Terre, Mtsamboro, Îlot Mtsamboro, chemin de crête, ♀ fr., 26.III.2002, Barthelat, Labarthe,Voynet-Fuasset, Dufour, M’Changama & Ali Sifari 796 (MAYOTTE, P!). — Bouzi [Îlot Bouzy], ster., IX.1850, Boivin s.n. (P!). — Îlot Bouzy, ♂ fl., 10.IV.2001, Hladik 6494 (K!, P!). — Îlot Bouzy, ♀ fr., 10.IV.2001, Hladik 6494bis (P!). — Îlot Bouzi, Mamoudzou, ♀ fl., 4.III.2002, Barthelat, Gomel & Ali Sifari 727 (G, K!, MAYOTTE, MO, P!). — Îlots Choazil, Malandzamiayatsini, sur la crête, 12°40’37”S, 45°03’28”E, ♂ fl., 29.IV.1999, Pignal, Pibot & Mas 1389 (P!). — Mont Hachiroungou, 12°42’43”S, 45°03’46”E, ♀ fl. and young fr., 16.IV.1996, Labat & Pascal 2744 (K!, MAYOTTE, MO, P!). — Plateau de Sohoa, ster., 5.IV.2001, Hladik, Barthelat, M’Changama & Sifary 6455 (P). — Sohoa, bord de plage de Mtsanga Nyamba, ♀ fl. & young fr., 5.IV.2001, Hladik, Barthelat, M’Changama & Sifary 6469 (K!, P!). — Sud, Mangrove d’Ironi, dans la pente en arrière de la mangrove, ♂ fl., 22.II.1975, Floret 1145 (P!). — Îles Comores, without further locality, ♀ fr., 1847-1852, Boivin s.n. (P!) GoogleMaps .
DESCRIPTION
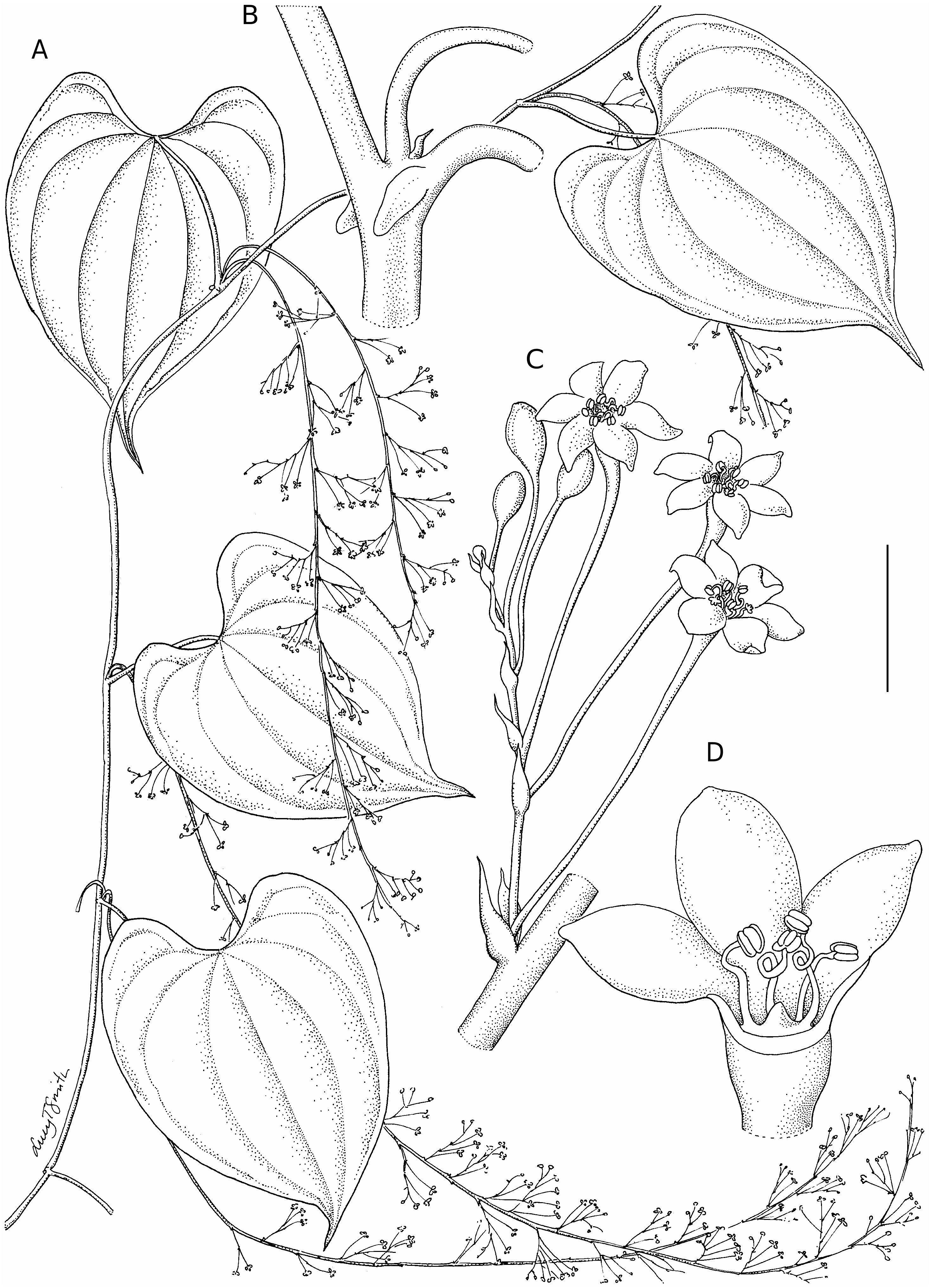
Twining vine to c. 10 m, stems annual from a fleshy tuber.Tubers annually replaced, those of current and previous rainy seasons usually present, divergent, buried 30-60 cm deep, to c. 50 × 3 cm, cylindric to cylindric-clavate, narrowing to crown, epidermis pale tan, parenchyma white, crown small, c. 0.5 cm in diameter, ovoid to dome-shaped, bearing thick white roots like stem nodes, tuber roots thinner, pale brown. Indumentum absent except on very young axillary vegetative buds and shoots, which have lax to dense, appressed, colourless, very rapidly caducous unicellular hairs to 0.2 mm long. Stems left-twining, to at least 5 mm in diameter, emerging from crown, white below ground with thick white roots at nodes, vertically oriented, above ground terete with weak longitudinal ridges, unarmed, cataphylls to 5 mm long present towards stem base. Leaves ( Fig. 2A View FIG ) alternate, blade 4.6-13.0 × 3.8-12.2 cm, ovate to broadly so or rarely orbicular, chartaceous, mid-green above, paler and grey-green below, veins 5 to the apex, with 1-2(-3) veins towards the base on both sides, margins entire, base deeply cordate, not lobed or auriculate, sinus 4.5-23 mm deep; apex 9-23 mm long, obtuse to rounded and acuminate with a 1-7 mm long, narrowly deltoid, brown to concolorous forerunner tip, primary venation prominent on lower leaf blade surface; petiole (1.5-) 2.4-7.3 cm long, strongly channeled on upper surface, lateral nodal spines (“stipules” of Burkill 1960) usually present ( Fig. 2B View FIG ), two fleshy, recurved, acute projections to c. 2 mm long on each side of amplexicaul petiole base, some collapsing on drying. Inflorescences one per axil, simple, racemose, pendent, sometimes with irregular warty galls. Male inflorescences ( Fig. 2A View FIG ) with a primary axis of (5-) 15-50 cm long or more (including 1.1-1.7 cm long peduncle), 0.5-1.5 mm wide (including wings), flattened and winged on both sides, nodes 4.5-14 mm apart, spacing greater towards inflorescence base, cymules ( Fig. 2C View FIG ) usually distichous, cymules of up to 7 flowers (first one or two branches of cymule very short), axis narrowed on the apical side of each node through the absence of its wings, cymule bracts two per node, 1-2 mm long, more or less equal in size, inserted at c. 90° to each other, ovate to elliptic, membranous with a slightly thickened midrib, apex acute to acuminate, first visible cymule branch to 0.8 mm long, all branches very slender. Female inflorescences ( Fig. 3A View FIG ) to at least 40 cm long, simple, spicate, flowers solitary, c. 10-30 per inflorescence, 5-20 mm apart, spacing greater towards base of axis as capsules develop but even and flowers on alternate sides of axis, axis flattened and winged like male inflorescence, width 1-2.3 mm, axis narrowed on the apical side of each node through the absence of its wings. Male flowers ( Fig. 2D View FIG ) pedicellate, scent not recorded, pedicels 1.4-9.2 mm long, longest in first flower of cymule (always at least 6 mm long when flower open), then decreasing in successive flowers of cymule, but never less than 4.3 mm and usually more than 5 mm long when the flower is open, very slender, only slightly thickened at apex, 0.2-0.5 mm wide; floral bract 0.5-1.3 × 0.2-0.5 mm, at pedicel base, elliptic to lanceolate, membranous, midrib scarcely thickened, base clasping cymule axis; tepal whorls not differentiated, inserted on a shallowly cup-shaped torus 0.4-0.8 × 0.5-1.0 mm, not constricted at the apex, walls relatively thick and opaque when dry; tepals 6, 0.8-1.5 × 0.5-1.0 mm, ovate to elliptic or oblong-elliptic, more or less patent to torus wall at anthesis, not usually fully reflexed, free or slightly fused at base, membranous, translucent, midrib scarcely thickened; stamens 6, erect, slightly exserted, appearing to be of irregular unequal length, filaments 0.3-0.9 mm long, free, filiform, contorted to coiled in the middle such that the anthers form a dense cluster at the torus mouth, anthers 0.15-0.3 × 0.15-0.4 mm, elliptic-oblong in outline, dorsifixed, pale yellow; pistillode a small, undifferentiated fleshy swelling at the centre of the torus. Female flowers ( Fig. 3B View FIG ) (sub)sessile until capsule development begins, scent not recorded; floral bract 1-1.8(-2.3) × 0.4-0.7 mm, narrowly ovate, long-acuminate, membranous with a thicker midrib, bracteole like bract but smaller, inserted at 90° to it and with a shorter acumen; ovary c. 3.4-7.4 mm long, 3-angled, narrowly elliptic to narrowly oblong in outline, becoming oblanceolate, narrowing at the apex into a ridged floral stipe 1.1-1.9 mm long, bearing a shallowly cup-shaped torus ( Fig. 3C, D View FIG ) 0.5-0.8 × 0.5-1.0 mm, dark and opaque when dry; tepals 6, not differentiated into two whorls, 0.7-1.2 × 0.5-0.8 mm, narrowly ovate to elliptic or elliptic-oblong, inserted on the torus rim, ascending to spreading, free or fused at the base, membranous, midrib scarcely differentiated, apex acute to obtuse; staminodia 6, 0.2-0.5 mm long, erect, length unequal, filamentodia straight to curved, antherodia not more than 0.1 mm long, styles 3, 0.7-1.1 mm long, fused for about half their length into an erect column, ascending to spreading above, stigmas narrowly ovate, deeply bifid, tips barely exserted from mouth of torus. Capsules ( Fig. 3E View FIG ) 23-37 × 13-18 mm, ascending at c. 45-30° to axis at dehiscence on a 3-5.5 mm long, narrowly conical, 3-angled pedicel which develops with the capsule, oblong to oblong-elliptic or sometimes broadening slightly towards apex, pale brown, darker towards wing margins, with chestnut brown flecking, base truncate to very shallowly cordate, apex acute to truncate, floral stipe persistent as an apiculus, splitting to give a narrower apiculus on each lobe of capsule, apical part only opening to release seed, seed wing basal and therefore concealed. Seeds ( Fig. 3F View FIG ) 6.5-10 × 4.5-8 mm excluding wing, lenticular-reniform to lenticular-ovoid, dull brown, wing 9-19 × 5-8.5 mm, orientated towards pedicel, oblong, membranous, golden-brown, translucent, apex obtuse to truncate.
REMARKS
The specimens Coulon 31 and 28 have the locality “Grande Comore, Îlot de Baudilé” on the label. However, Grande Comore does not have îlots (it is geologically different from Mayotte) and the locality name is close in spelling to Bandrélé, so we believe that they are most likely to have been collected at the Îlot de Bandrélé off the coast of Mayotte.
Dioscorea mayottensis can be recognized in the vegetative state by its rapidly caducous pubescence on very young buds only. Dioscorea comorensis is pubescent on buds, young shoots and around the axils; its hairs are more persistent. When in flower, the male plant has a very distinctive inflorescence arrangement, with alternate cymules of (4-)5-7 flowers 4.5-14 mm apart. In D. comorensis , the male inflorescence has irregular to subopposite cymules of 3(-4) flowers 0.7-5.0(-7.3) mm apart. The cymule branches and pedicels of D. mayottensis are more slender throughout than those of D. comorensis , and the pedicels are at least 6 mm long in the first flower of the cymule, usually more than 5 mm long and never less than 4.3 mm long in the subsequent flowers at anthesis. The pedicels of the male flowers of D. comorensis do not exceed 3.1 mm long. The torus size and shape differs in the two species (0.4-0.8 × 0.5-1 mm in D. mayottensis , 0.2-0.5 × 0.8-1.5 mm in D. comorensis ). Finally, the filaments of D. mayottensis are coiled or contorted rather than reflexed. The numerous differences in reproductive morphological characters suggest that D. mayottensis and D. comorensis have different pollinators and thus are reproductively as well as spatially isolated. Female plants of D. mayottensis can be identified by characters including the winged inflorescence axis, the even spacing and alternate positioning of the ovaries and the 1.1-1.9 mm long floral stipe (see Fig. 3B View FIG ) at the ovary apex (0.4-0.8 mm long in D. comorensis ). The capsules and seeds of D. mayottensis appear to be larger than those of D. comorensis but this may reflect the state of development of the capsules of Labat et al. 3717, the only female specimen of the latter.
DISTRIBUTION AND ECOLOGY
Endemic to Mayotte. Littoral vegetation, usually in dry forest and scrub to dense humid forest from near sea level to 450 m in elevation, often on slopes. Dioscorea mayottensis occurs on several substrates, including seasands, calcareous, cineritic and lateritic soils (but not when the latter are compacted). It has been found in abandoned cultivated areas, so it can tolerate a degree of disturbance, and can be encountered in active cultivation as well as in managed wild populations.
VERNACULAR NAMES
Mitrou (Shimaoré), Oviala (Shibushi).
USES
Tubers edible, said to have a very good flavour.
CONSERVATION STATUS
IUCN Red List category NT ( IUCN 2001). There are c. 11 known populations on Mayotte based on the herbarium specimens cited above. Both deforestation and over-utilisation are probable threats. Dioscorea mayottensis is legally protected in Mayotte under arrêté préfectoral DAF no. 42-2006, article 2 of which forbids the sale, purchase or placing for sale any part of the wild plant; it may be collected for consumption only. Nevertheless, its desirability as a food makes a study of its sustainable use a priority to ensure its long-term survival.
INFLORESCENCE MORPHOLOGY
IN D. COMORENSIS AND D. MAYOTTENSIS
Both D. comorensis View in CoL and D. mayottensis View in CoL possess a cymule in the axil of a bract and bracteole at each node of the inflorescence axis. Burkill (1960) regarded the switch from a single flower on a raceme or spike to a cymule as a means of maximising male reproductive investment. Such cymulose inflorescences were called drepania by Weberling (1989). Evidence that drepania have evolved from racemose and spicate partial inflorescences is found in the bract and/or bracteole at base of each cymule, which are homologous with the floral bract and bracteole. The phylogeny obtained by Wilkin et al. (2005) suggests that this change has occurred many times in the genus Dioscorea View in CoL across its geographical range and in most of the main subclades. As Figure 2C View FIG shows, D. mayottensis View in CoL appears to have two cymule bracts of more or less equal size. However, the second bract may be the floral bract of the first flower of the cymule, which has its pedicel inserted just above the base of the cymule axis and always has the longest pedicel in the cymule. The length of the inflorescence and the number of flowers in each cymule makes the male inflorescence of D. mayottensis View in CoL highly floriferous. Malagasy endemic species with drepania include D. sterilis Weber & Wilkin View in CoL ( Weber et al. 2005), D. maciba Jum. & H.Perrier View in CoL ( Burkill & Perrier de la Bâthie 1950), D. sambiranenisis R.Knuth ( Burkill & Perrier de la Bâthie 1950), D. namorokensis Wilkin View in CoL ( Wilkin et al. 2002), D. bemarivensis Jum. & H.Perrier View in CoL ( Haigh et al. 2005) and D. arcuatinervis Hochr. View in CoL ( Burkill & Perrier de la Bâthie 1950). Both the phylogeny in Wilkin et al. (2005) and the diversity of other morphological characters in these taxa indicate that there has been more than one origin of drepania in Madagascar alone.
RELATIONSHIPS OF D. COMORENSIS View in CoL AND D. MAYOTTENSIS View in CoL WITH YAMS FROM MADAGASCAR
The left twining stems, basally winged seeds and lateral nodal organs of both D. comorensis and D. mayottensis and the tuber morphology of the latter suggest that they are most closely related to species from Madagascar rather than Africa. The modification of the torus and long inflorescences with winged axes suggest links to the campanulate-flowered species group in Madagascar, which includes D. seriflora Jum. & H.Perrier from the humid East and D. maciba Jum. & H.Perrier from the seasonally dry West. However, only D. maciba of that species group possesses flowers in cymules. A well-sampled phylogenetic study of the Malagasy clade of Dioscorea ( Wilkin et al. 2005) is needed to test this hypothesis.
KEY TO THE ENDEMIC AND INTRODUCED YAMS ( DIOSCOREa View in CoL L.) OF THE COMORO ARCHIPELAGO BASED ON VEGETATIVE CHARACTERS AND DISTRIBUTION
1. Leaves compound ....................................................................................................... 2 — Leaves entire or rarely lobed ....................................................................................... 3
2. Tubers ovoid to fusiform, several in a dense cluster. Hairs quite coarse, often dense, especially on leaf lower surface ..................................................................... D. quartiniana
— Tubers long, terete, spreading horizontally, with swollen apices. Hairs sparse, also with flattened, deltoid hairs mainly on the leaflet margins ............................ D. bemarivensis
3. Tuber globose with a flattened base and depressed apex. Mature leaves with acuminate, thickened apices with inrolled margins, juvenile leaves lobed ................ D. sansibarensis
— Tuber(s) not as above. Leaf apex if acuminate not thickened or with inrolled margins, margins always entire ................................................................................................. 4
4. Leaves opposite throughout. Stems right twining ....................................................... 5 — Leaves alternate at least on distal shoots. Stems left twining ........................................ 6 5. Stems square in cross-section, with 4 longitudinal wings ................................. D. alata — Stems terete, wingless ........... D. minutiflora (sensu Burkill & Perrier de la Bâthie 1950)
6. Hairs persistent, T-shaped, usually dense and greyish. Stem spiny .............. D. esculenta — Hairs absent or caducous, never T-shaped. Stem usually unarmed .............................. 7
7. Two membranous semicircular projections at petiole base/node. Axillary bulbils usually abundant ................................................................................................... D. bulbifera
— Two fleshy, recurved, acute projections usually present at petiole base/node. Axillary bulbils absent .................................................................................................................. 8
8. Hairs absent except on very young axillary vegetative buds and shoots, which have lax to dense, appressed, very rapidly caducous hairs to 0.2 mm long. Mayotte........................ .................................................................................................. D. mayottensis sp. nov.
— Hairs caducous (but more persistent than those of D. mayottensis ), to 0.7 mm long, dense on young vegetative buds and usually so on shoots, sparsely persistent around the axil and on the leaf blade lower surface, especially on the veins near the point of petiole insertion, usually appressed but sometimes erect on buds. Mwali & Njazidja .......... D. comorensis


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dioscorea mayottensis Wilkin
| Wilkin, Paul, Hladik, Annette, Labat, Jean-Noël & Barthelat, Fabien 2007 |
D. mayottensis
| Wilkin & Hladik & Labat & Barthelat 2007 |
D. mayottensis
| Wilkin & Hladik & Labat & Barthelat 2007 |
D. mayottensis
| Wilkin & Hladik & Labat & Barthelat 2007 |
D. MAYOTTENSIS
| Wilkin & Hladik & Labat & Barthelat 2007 |