
Chondrocyclus kevincolei, Cole, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.569 |
|
publication LSID |
lsid:zoobank.org:pub:79BE13FC-B840-4C39-8D25-3328BDCC44D2 |
|
persistent identifier |
https://treatment.plazi.org/id/2C3567A2-1004-4E9B-9452-C6E657323E18 |
|
taxon LSID |
lsid:zoobank.org:act:2C3567A2-1004-4E9B-9452-C6E657323E18 |
|
treatment provided by |
Plazi |
|
scientific name |
Chondrocyclus kevincolei |
| status |
sp. nov. |
Chondrocyclus kevincolei View in CoL sp. nov.
urn:lsid:zoobank.org:act:2C3567A2-1004-4E9B-9452-C6E657323E18
Figs 5 View Fig , 10 View Fig P–Q, 26
Chondrocyclus convexiusculus – Connolly 1939: 537 View in CoL .
Diagnosis
Shell small, depressed, lenticular; periostracum with axial costae producing spiral rows of simple hairs; spiral grooves on shell corresponding with rows of hairs; operculum duplex, exterior portion very shallowly concave, with step-shaped multispiral lamella terminating in a short fringe with uneven edge, diameter of exterior portion less than diameter of inner disc; radula with three large cusps on second lateral tooth.
Etymology
Named for the author’s husband, Kevin Cole, in acknowledgement of his indispensable assistance on fieldtrips.
Type material examined
Holotype
SOUTH AFRICA – Western Cape • Platbos Forest near Gansbaai, Western Cape Milkwood Forest with large trees including Sideroxylon inerme Linne and Celtis africana N.L. Burm ; 34.5671°S, 19.4495°E; 13 Aug. 2014; M. and K. Cole leg.; in leaf litter; NMSA W9270/T3073 . ( Fig. 10 View Fig P–Q) GoogleMaps
Paratypes
SOUTH AFRICA – Western Cape • 1 specimen; same collection data as for holotype; ELM D18004/ T 85 • 2 specimens; same collection data as for holotype; ELM W 3898 View Materials / T 86 • 327 specimens; same collection data as for preceding; 9 Sep. 2009; M. Cole, K. Cole, D-J Hodgkinson and T. Pretorius leg.; ELM D16921/ T 81 • 90 specimens; same collection data as for preceding; ELM W 3615 View Materials / T 82 • 20 specimens; same collection data as for preceding; NHMUK 20120282 About NHMUK • 23 specimens; same collection data as for preceding; NMSA P 0598 About NMSA / T 4121 • 18 specimens; same collection data as for preceding; NMW. Z.2012.065.00010 • 15 specimens; same collection data as for preceding; RMNH MOL.330503 • 47 specimens; same collection data as for preceding; 18 Apr. 2012; M. Cole, D. Herbert and L. Davis leg.; ELM D16998/ T 83 • 5 specimens; same collection data as for preceding; ELM W 3659 View Materials / T 84 • 27 specimens; Bredasdorp, about 5 miles to SW, on Elim Rd ; 34.5450° S, 19.9614° E; ex Transvaal Museum; dry bush on hillside; NMSA W 575 About NMSA / T 4123 GoogleMaps • 34 specimens; Bredasdorp, 7 km south west on R 43 to Elim , bushy valley with dam; 34.5912° S, 19.9992° E; 35 m a.s.l.; 13 Aug. 2014; M. Cole leg.; ELM D17983/ T 87 GoogleMaps • 15 specimens; same collection data as for preceding; ELM W 3890 View Materials / T 88 GoogleMaps • 23 specimens; Grootbos Nat. Res., Gansbaai area, Milkwood forest ; 34.5421° S, 19.4153° E; 7 Oct. 2007; D. Herbert and L. Davis leg.; in leaf-litter; NMSA W 5659 About NMSA / T 4122 GoogleMaps • 45 specimens; Grootbos Private Nature Reserve, between Stanford and Gansbaai, Western Cape Milkwood Forest ; 34.5419° S, 19.4122° E; 203 m a.s.l; 13 Aug. 2014; M. Cole leg.; D18005/ T 89 GoogleMaps • 6 specimens; Gansbaai dune scrub; 34.5795° S, 19.3442° E; 13 Feb. 2005; A. Moussalli and D. Stuart-Fox leg.; in leaf-litter; NMSA W 3537 About NMSA / T 4124 GoogleMaps .
Description
SHELL ( Fig. 26 View Fig A–F). Small, depressed, lenticular, adult diameter 3.59–5.34 mm, height 1.59–2.84 mm, diameter:height 1.68–2.37 (n = 87 from 4 populations; variations in dimensions between populations are given in Table 4 View Table 4 ). Spire little exserted, protoconch large and mammillate ( Fig. 26A View Fig ). Embryonic shell ( Fig. 26G View Fig ) approx. 2.25 whorls, almost smooth but microscopically malleate at tip, junction between embryonic shell and teleoconch evident with development of costae on teleoconch. Teleoconch comprising just over two whorls, very convex, rapidly increasing, suture deeply impressed. Aperture circular, last whorl descending near aperture, peristome simple, continuous and free. Umbilicus wide and deep, exposing all the whorls Fig. 26C, F View Fig ). Periostracum glossy and lacquer-like with lamellate axial costae at regular intervals on last whorl, 51–75 (n = 22) in Platbos population, but vary between populations ( Table 4 View Table 4 ), which produce spiral rows of simple hairs varying in different populations from approx. 4–14 on last whorl, longest at periphery and shortest around umbilicus; intervals between axial costae with approx. eight microscopic axial threads ( Fig. 26H View Fig ); shell bears spiral grooves corresponding with the rows of hairs ( Fig. 26 View Fig D–F). Shell translucent when fresh; two colour morphs present reddish brown and creamy white.
LIVING ANIMAL. Varies in colour between populations from creamy white with slight pigmentation on tentacles to very dark grey (see below).
OPERCULUM ( Fig. 26 View Fig J–K). Duplex, shallowly concave; multispiral lamella of outer portion with 4.5 whorls, each step-shaped, growing edge of lamella at angle, perpendicular where it is attached to disc; fringe very short and solid, with a frayed distal edge even in fresh specimens, fringe of each whorl not fused to lamellar blade of following whorl, diameter of outer multispiral portion smaller than diameter of inner disc due to very short fringe; operculum retractile.
RADULA ( Fig. 26I View Fig ). Rachidian with five cusps, middle one longer than two cusps on either side of it; first and second lateral teeth each with four cusps and a vestigial 5 th, the third cusp from centre the largest.
PENIS ( Fig. 26L View Fig ). Shaft more or less cylindrical and slightly flattened dorsoventrally, distal half slightly expanded on left side, numerous annular rugae, distal end smooth, intromittent organ short.
Distribution and habitat
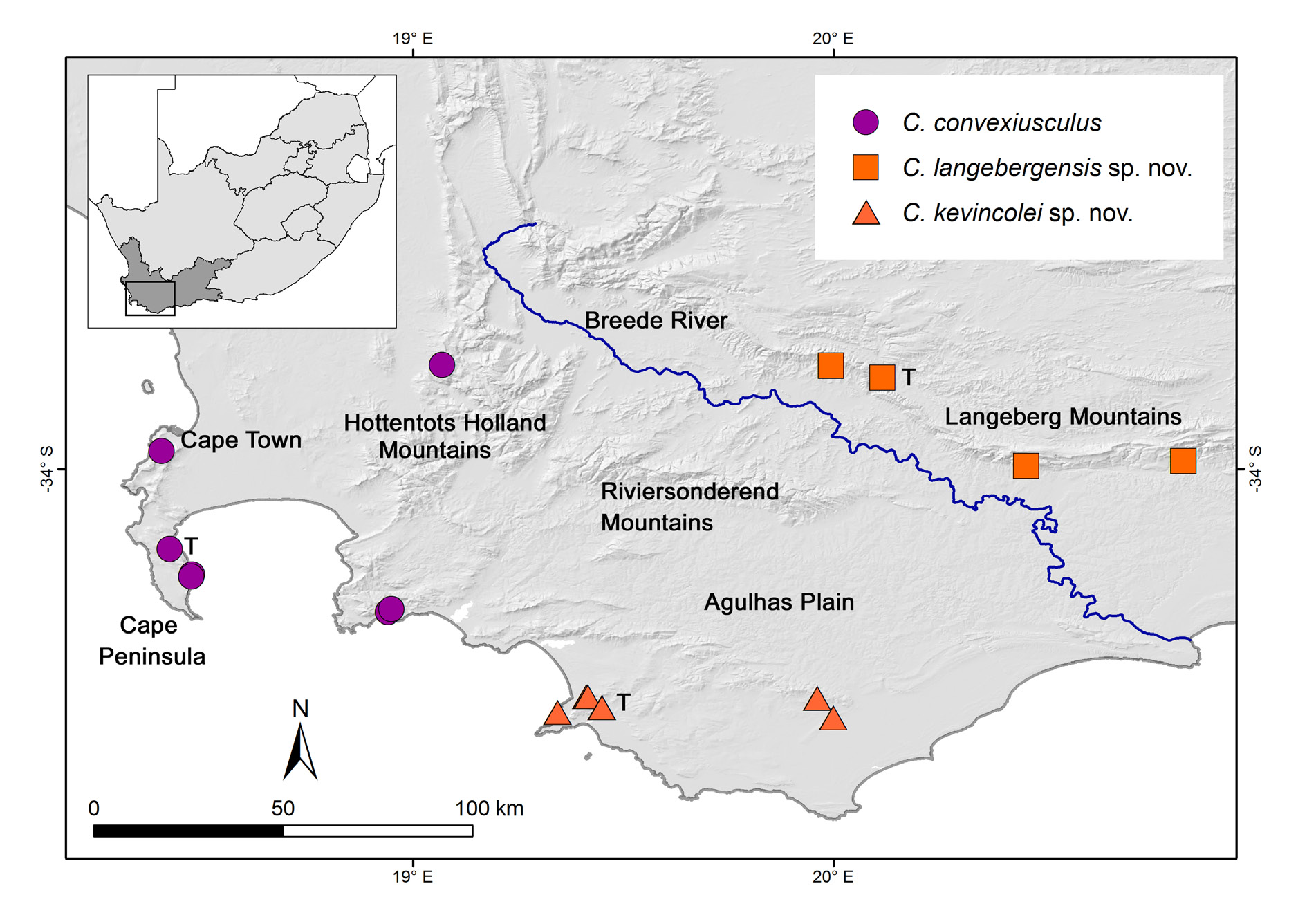
Western Cape, Agulhas Plain, between Gansbaai and Bredasdorp. Various forested or bushy habitats in patches of Western Cape Milkwood Forest (part of Southern Coastal forest group) ( von Maltitz et al. 2003) with large trees, characterised by white milkwoods ( Sideroxylon inerme ), white stinkwoods ( Celtis africana ) and white pear ( Apodytes dimidiata E. Mey. ex Arn. ), including the southernmost forests in Africa (Platbos and Grootbos), dune scrub at Gansbaai and ‘dry bush on hillside’ (south-west of Bredasdorp); in leaf-litter ( Fig. 5 View Fig ).
Remarks
The distinct spiral grooves on the shell of C. kevincolei sp. nov. ( Fig. 26 View Fig D–F) are unique among species of Chondrocyclus although there are sometimes faint traces of spiral grooves on shells of A. isipingoensis gen. et comb. nov. and C. langebergensis sp. nov. visible only at very high magnification. Chondrocyclus langebergensis sp. nov. differs from C. kevincolei sp. nov. in its lack of prominent spiral grooves on the shell, the spiral rows of long hairs are concentrated around the periphery, the protoconch is more strongly sculptured and the opercula differ. In C. langebergensis sp. nov., the lamellar fringe of each spiral whorl of the operculum emanates from a ridge at the base of the lamellar blade and is fused to the lamellar blade of the following whorl. The terminal fringe is long and overlaps the base when viewed from above ( Fig. 25F View Fig ). In C. kevincolei sp. nov. the fringe emanates from near the top of the lamellar blade and is very short and appears distinct from the following lamellar whorl; the outer saucershaped portion is smaller than the polished base ( Fig. 26J View Fig ). These differences together with molecular evidence, are considered to warrant recognition of at least two distinct species, one in the Langeberg Mountains and the other on the relatively low-lying coastal region of the Agulhas Plain. The Langeberge and Agulhas Plain are separated by the Breede River valley and by a belt of relatively arid vegetation, consisting mainly of Ruens Shale Renosterveld and Agulhas Limestone Fynbos ( Rebelo et al. 2006), which developed in the Pliocene ( Cowling et al. 2008). Molecular studies in unrelated faunal taxa also provide evidence for separate lineages in the Langeberge and the Agulhas Plain (e.g., Price et al. 2007; Moussalli et al. 2009; Gouws et al. 2010; McDonald & Daniels 2012) and for an absence of gene flow between these areas ( Myburgh & Daniels 2015). Species of Chondrocyclus have not been recorded in the Riviersonderendberge where apparently suitable habitat exists.
There are morphological differences between populations of C. kevincolei sp. nov. ( Table 4 View Table 4 ). The number of spiral rows of hairs is fewer in the Bredasdorp population (only 4–5 as opposed to 8–15 in the Platbos and Grootbos populations). The colour of the living animal is creamy white with only slight pigmentation of the tentacles in the Platbos population and dark grey to almost black at Bredasdorp. There were relatively large genetic divergences between populations ( Cole et al. 2019). Their long branch lengths ( Fig. 1 View Fig ) suggest isolation in shrinking refuges as arid-adapted vegetation replaced forest (see Tolley et al. 2006, 2008). Recent radiations of faunal taxa in this region are associated with adaptation to open habitats ( Tolley et al. 2006; Herbert & Moussalli 2010). Sea level changes have also had dramatic effects on vegetation of the Agulhas Plain ( Linder 2003) and its biota with marine transgressions potentially restricting ancestral populations of Chondrocyclus to refuges at higher elevations and thus driving allopatric divergence or eliminating closely related taxa at lower elevations. Further studies including unsampled populations on the Agulhas Plain and additional markers may shed light on whether separate populations may warrant species status. It was, however, decided to treat these as one species pending further investigation. By contrast, C. langebergensis sp. nov. occurs in relatively stable relictual forest patches in the Langeberge and has undergone little change.
Genus Afrocyclus gen. nov.
urn:lsid:zoobank.org:act:0C542886-FB1A-4550-BBC2-80BBD28FA01E
Type species
Afrocyclus isipingoensis ( Sturany, 1898) View in CoL gen. et comb. nov.
Diagnosis
Shell dextral, small, depressed; aperture circular, last whorl descending near aperture, peristome not thickened, continuous and free; umbilicus wide and deep, exposing all the whorls; periostracum glossy and lacquer-like with lamellate axial costae at regular intervals, usually expanded into spiral rows of hairs; operculum duplex, corneous; inner portion consists of a thin disc which grows outwards in a tight spiral, smooth on inside where it attaches to foot; fused to disc on its outer surface is a multispiral, elevated horny blade-like lamella formed by fused bristles; a fringe of fused bristles emanates from outer surface of each lamellar whorl; radula taenioglossate, two large cusps on second lateral tooth; penis lies dorsally immediately behind right tentacle and consists of a stout, muscular, cylindrical shaft, wrinkled along most of its length due to annular rugae, and a terminal, tapering intromittent organ without flagellum; seminal tube completely enclosed without seminal groove.
Etymology
The specific name is derived from ‘Africa’ and refers to its Afromontane occurrence.
Remarks
mtDNA data clarified that Afrocyclus gen. nov. constituted a radiation distinct from that of Chondrocyclus s.s. ( Cole et al. 2019). Specimens of Afrocyclus gen. nov. can in most cases be recognised at a glance on morphological grounds, being very small, very depressed (the apex can even be concave), discoid shells with spiral rows of hairs projecting from the axial costae. This morphotype is widely distributed in latitude and altitude, occupying isolated Afromontane forest patches along the Great Escarpment from northern South Africa (Limpopo province, 23° S) to the Eastern Cape (32.7°S) and extending to the coast ( Fig. 2 View Fig ). mtDNA data showed that what was previously lumped as Afrocyclus isipingoensis gen. et comb. nov. is a species complex, and within it, external morphology is relatively conserved, making it difficult to distinguish taxa by morphology alone. A. exsertus gen. et comb. nov., which occupies a narrow zone on the east coast, is strikingly different morphologically, bearing no hairs and with the axial lamellae only slightly expanded around the periphery. The operculum of the latter also deviates dramatically from the flat, fragile operculum of most specimens of Afrocyclus gen. nov. and is more robust with a rigid fringe consisting of bunches of short bristles. Phylogenetic relationships within Afrocyclus gen. nov. are not fully resolved and further molecular and morphological data from throughout its range are required to revise this complex.
Key to species of Afrocyclus View in CoL gen. nov.
1. Axial lamellae slightly expanded at periphery, but not produced into hairs........................................ ............................................................... A. exsertus ( Melvill & Ponsonby, 1903) View in CoL gen. et comb. nov.
– Axial lamellae expanded into multiple spiral rows of hairs .............................................................. 2
2. Spiral rows of periostracal hairs include a row of very short hairs immediately below suture on body whorl .................................................................. A. isipingoensis ( Sturany, 1898) View in CoL gen. et comb. nov.
– No spiral row of very short periostracal hairs immediately below suture......................................... 3
3. Three spiral rows of hairs at periphery; upper edge of rachidian tooth smooth .................................. .................................................................................................................. A. oxygala View in CoL gen. et sp. nov.
– More than three spiral rows of hairs on body whorl; upper edge of rachidian tooth serrated........... 4
4. Protoconch strongly malleate ...................................................................... A. potteri View in CoL gen. et sp. nov. – Protoconch malleate, but not strongly .......................................................... A. bhaca View in CoL gen. et sp. nov.





| NMSA |
KwaZulu-Natal Museum |
| ELM |
East London Museum |
| T |
Tavera, Department of Geology and Geophysics |
| W |
Naturhistorisches Museum Wien |
| M |
Botanische Staatssammlung M�nchen |
| K |
Royal Botanic Gardens |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| NMW |
Naturhistorisches Museum, Wien |
| Z |
Universit�t Z�rich |
| RMNH |
National Museum of Natural History, Naturalis |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| S |
Department of Botany, Swedish Museum of Natural History |
| E |
Royal Botanic Garden Edinburgh |
| R |
Departamento de Geologia, Universidad de Chile |
| A |
Harvard University - Arnold Arboretum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Caenogastropoda |
|
Order |
|
|
Family |
|
|
Genus |
Chondrocyclus kevincolei
| Cole, Mary L. 2019 |
Chondrocyclus convexiusculus – Connolly 1939: 537
| Connolly M. 1939: 537 |