
Madenemura andringitrensis Paulian, 1959
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4059.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:9A0F1819-753E-471E-81F3-06A9719223D8 |
|
DOI |
https://doi.org/10.5281/zenodo.6100640 |
|
persistent identifier |
https://treatment.plazi.org/id/0E0C0B57-FFF6-FFDD-2BBC-27E0F94DFCE6 |
|
treatment provided by |
Plazi |
|
scientific name |
Madenemura andringitrensis Paulian, 1959 |
| status |
|
Madenemura andringitrensis Paulian, 1959 View in CoL
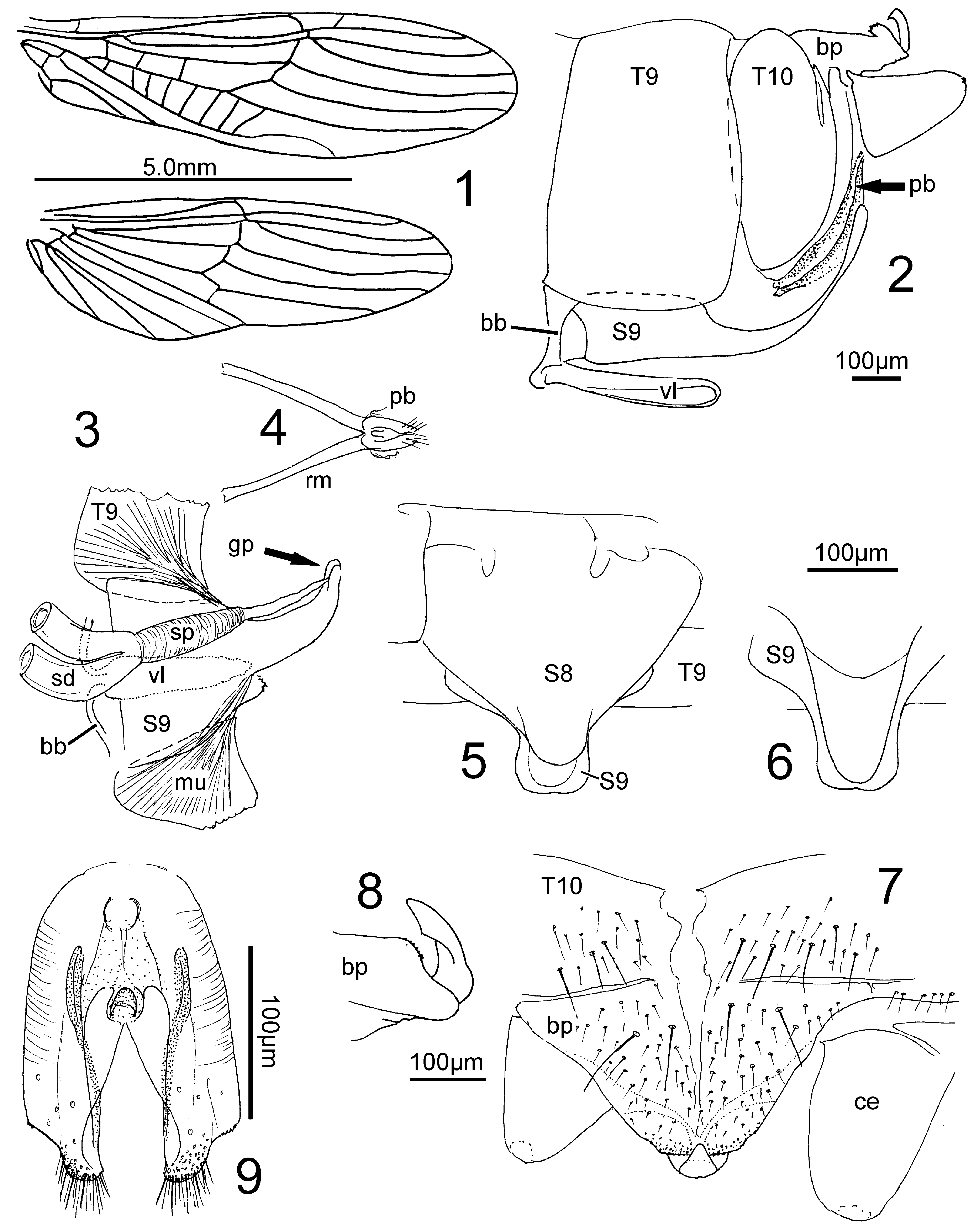
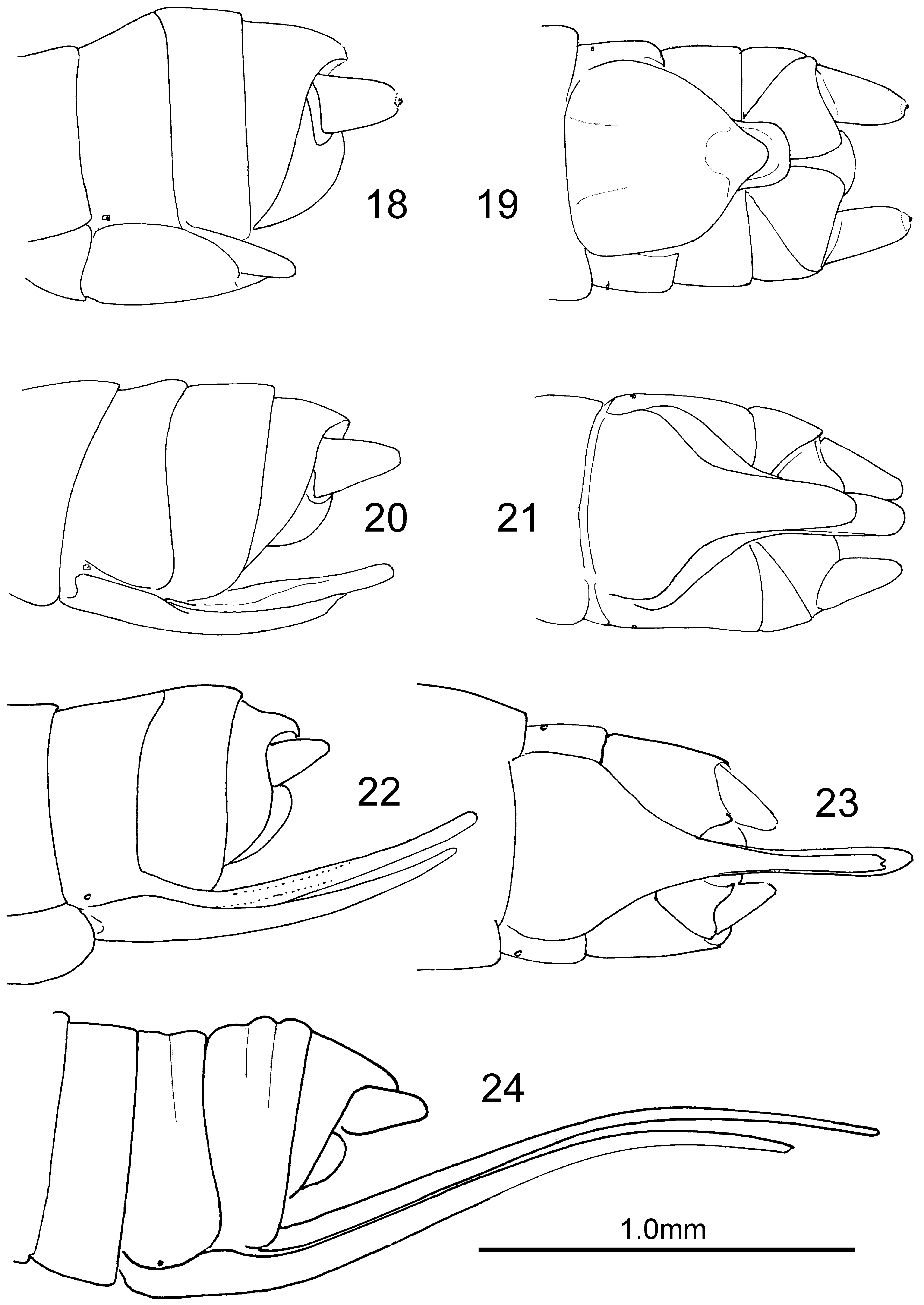
( Figs. 3–9 View FIGURES 1 – 9 , 18, 19 View FIGURES 18 – 24 , 25, 26 View FIGURES 25, 26 )
= Madanemura perrieri Paulian, 1959 , new synonymy. = Madanemura sp. 2 ♂ of Paulian (1959).
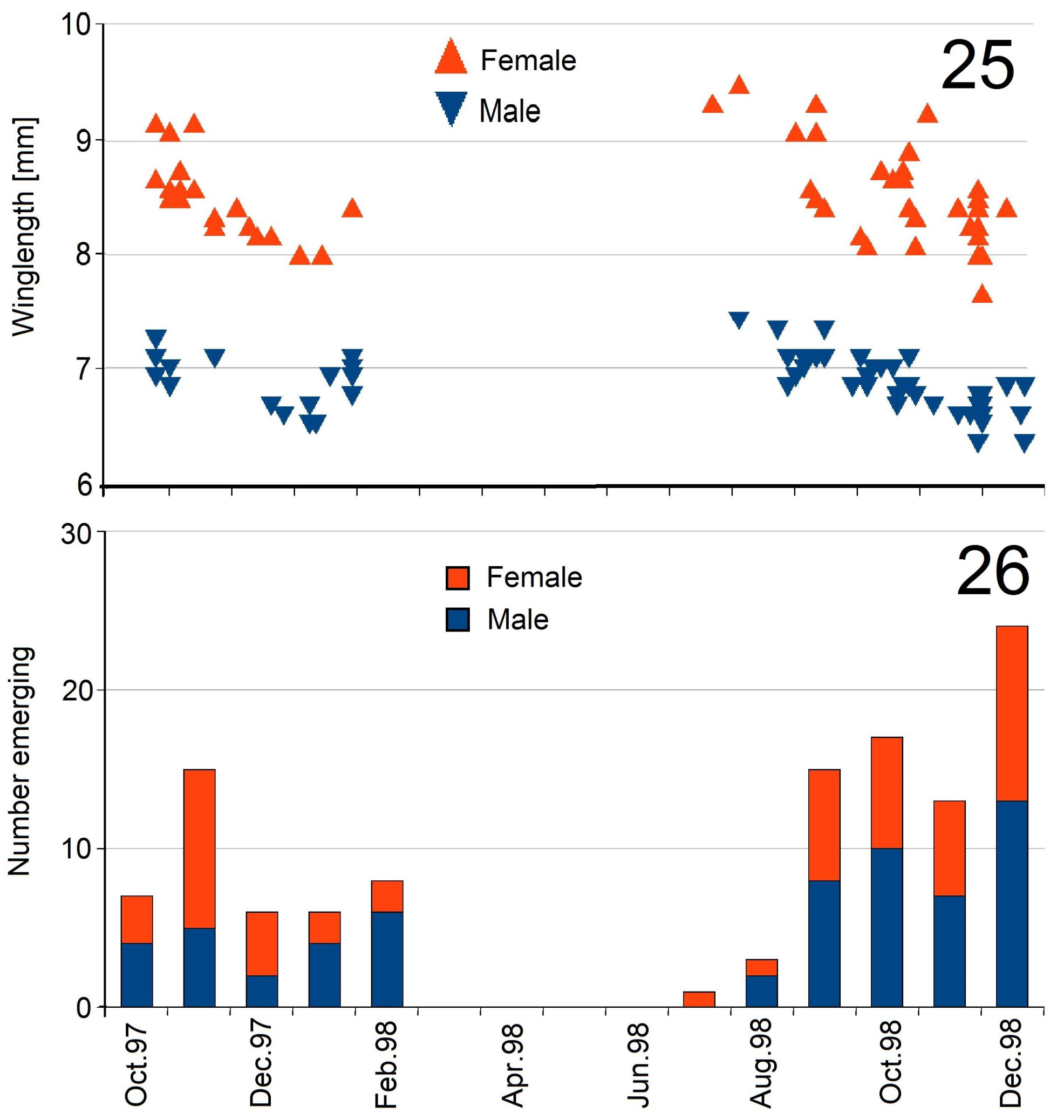
Material studied. Madagascar, Ranomafana National Pk., Mariavaratra stream, 2115.629'S, 4725.149'E; emergence trap, J. Benstead, 1997-1998, 61♂♂, 54♀♀; the seasonal distribution is shown in Fig. 26 View FIGURES 25, 26 . Prov. Fianarantsoa, 7km W Ranomanfana, 900m, W.E.Steiner (~ 22.223°S, 46.929°E; all USNM): Malaise trap in small clearing, 20– 31.01 .1990, 2♂, 1♀; 01.– 07.03.1990, 1♂; at black light: 20– 31.01.1990, 1♀; 17.– 22.02.1990, 1♂, 1♀; 01.– 0 7.03.1990, 1♂; Malaise trap across stream montane rain forest, 20.– 24.03.1990, 3♀; 08.– 13.03.1990, 1♀.
Larvae, probably M. andringitrensis : Madagascar, Ranomafana National Pk., Mariavaratra stream, 2115.629'S, 4725.149'E, J.P.Benstead, 8-19-98, 15 larvae; Madagascar, Ranomafana N.P. E4725.458' S2116.040' 26 Oct 1998 B, 3 larvae; Madagascar, Ranomafana N.P. 4725.149'E 2115.629'S, 26 Oct 1998, A, 3 larvae; Tolongoina stream, nr. Ranomafana N. P. Madagascar S2115.417' E4727.440' 7-2-98 J.P.Benstead, 1 last instar larva; Fompohonona stream, Ranomafana N.P., Madagascar S2116.040' E4725.458' 6-15-98 J.P. Benstead, ca. 10 larvae; same site, 8- 19-98 J.P. Benstead, 4 larvae; Vatoharanana stream, Ranomafana N.P., Madagascar S2117.407' E4725.677' 6-29-98 J.P. Benstead, 2 larvae (all USNM).
Habitus. As for the genus; WL of males 6.3–7.4mm, of females 7.7–9.5mm. Wing length varies with time ( Fig. 25 View FIGURES 25, 26 ).
Male. Tergite 10 longitudinally divided by a pale hairless median zone which is wide in front and narrows as it continues onto the baseplate of the epiproct where it tapers. Baseplate of epiproct separated from T10 by a narrow transverse pale and hairless line. The plate is almost parabolic, wide in front, narrowing backwards, a transverse strip in front of the short truncate caudal edge is beset with small black spinules. The sclerites supporting the epiproct originate on the underside of the baseplate, in cleared specimens their edges can be seen by transparency. Together they form the wide peduncle of the narrow erect hook-like part of the epiproct. Its apex is triangular, with blunt tip ( Fig. 7 View FIGURES 1 – 9 ). In side view the spinulose area of the baseplate appears as a shallow bulge over which the epiproct curves ( Fig. 8 View FIGURES 1 – 9 ).
The paraproct body ( Fig. 9 View FIGURES 1 – 9 ) is stout, about twice as long as wide and about half as long as the ventral lobe of S9. Its rounded front end includes a median sclerotised structure which in side view appears like a blunt, forwardcurved tube. The sides of the paraproct are elongate lobes with large circular sensilla, mainly on the outside. The apical portion bears numerous straight setae. Along the ventral edge (lying medially in slide mounts) is an elongate sclerite which is wide in the basal third, but caudally grows narrower and curves towards the medial edge.
Female. The ovipositor is very short, extending only to the base of the paraprocts ( Figs. 5, 6 View FIGURES 1 – 9 ). S8 is caudally extended, triangular, slightly raised along the midline, the end soft and projecting as a blunt knob. The wide, medially excised base of S9 is concealed by S8, caudally it projects as a broad flat process with a central soft pale area surrounded by a fairly wide sclerite with fine pilosity. The postgenital plate extends a little beyond the short subgenital plate of S8.
Notes. I recognize the species by the unusually short and wide ovipositor which looks “fleshy”, a term used by Paulian (1959) only in the description of M. andringitrensis . The small illustration of M. sp. 2 ♂ of Paulian (1959) fits the present males. Eighty-nine percent of the emergence trap catches belong to this species, the sex rate is balanced (53% males; Fig. 26 View FIGURES 25, 26 ), the conspecificity of sexes is beyond doubt. See the notes on Ecology below.
Paulian (1959) apparently noticed some terminal annulation around the rudimentary end segment of the female cercus and considered it was plurisegmented. However, such subterminal annulation occurs occasionally in many Nemouroidea and concerns only the pilosity. This annulation and differences in the posture of the ovipositor were used to distinguish M. perrieri which I regard as conspecific. The nominal taxa in question were all taken in the same geographic area, roughly between 21.26°S / 47.42°E and 22.22°S / 46.93°E.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Madenemura andringitrensis Paulian, 1959
| Zwick, Peter 2015 |
Madanemura perrieri
| Paulian 1959 |