
Madenemura Paulian, 1949
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4059.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:9A0F1819-753E-471E-81F3-06A9719223D8 |
|
DOI |
https://doi.org/10.5281/zenodo.6100634 |
|
persistent identifier |
https://treatment.plazi.org/id/0E0C0B57-FFF4-FFD9-2BBC-220FF982FD13 |
|
treatment provided by |
Plazi |
|
scientific name |
Madenemura Paulian, 1949 |
| status |
|
Genus Madenemura Paulian, 1949 View in CoL
Type species: M. capuroni Paulian, 1949 (by monotypy).
= Tsaranemura Paulian, 1951 View in CoL ; type species: T. culminalis Paulian, 1951 View in CoL , by monotypy. New synonymy. = Madanemura Paulian, 1959 (lapsus).
Species can be identified only by examining genitalia, the other distinguishing characters proposed by Paulian vary with degree of maturity, body posture, and condition at or after preservation. There was no decernable individual variation in genitalia.
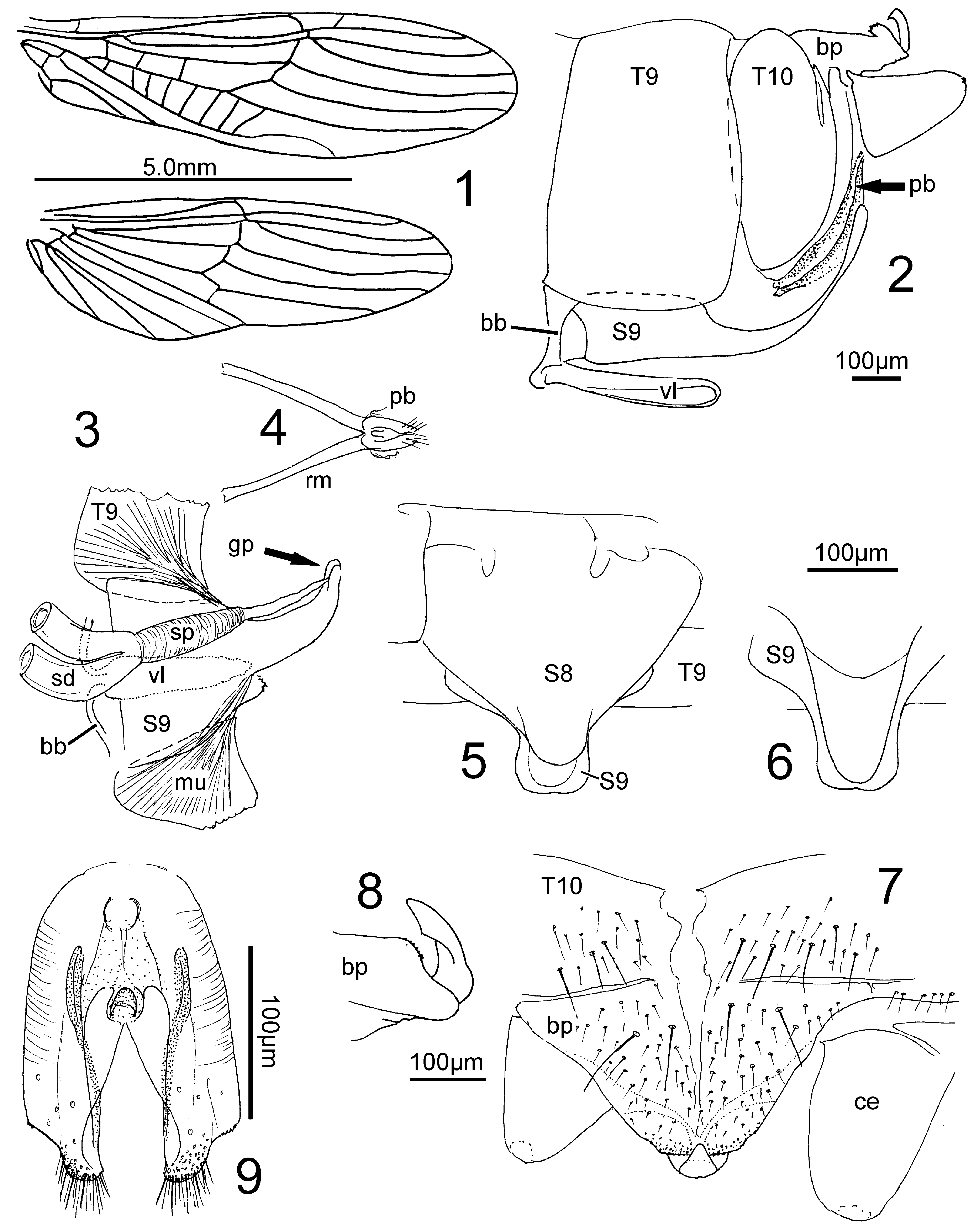
Habitus. Resembling slender Nemouridae View in CoL , except that the long wings are longitudinally folded around the body instead of resting flat above the abdomen. Head with the large eyes much wider than the pronotum, ocelli unmodified. Antennae long and thin, the 5-segmented maxillary palpus is curved back in a semicircle, the 2- segmented labial palpus is unmodified. The legs are slender, the tip of the hind femur extends back to near the abdominal tip. Wings long and narrow, front wing with 2 crossveins between M and Cu1, and ca 5 between Cu1 and Cu2. A1 straight, A2+3 with common base. No crossveins in hindwing, 5 straight anal veins ( Fig. 1 View FIGURES 1 – 9 ). Abdomen membranous, except small anterolateral sclerites, and the terminal segments. Cercus one-segmented, with wart-like terminal rudiment.
Uniformly brownish, also the wings. The base of the antennal flagellum may be lighter than the rest of the antenna. The legs are yellowish with dark pattern which is most expressed on the long hind leg: femur dark at base, on a middle patch, and near the knee. Tibia dark in subgenal area, tip and tarsus infuscate.
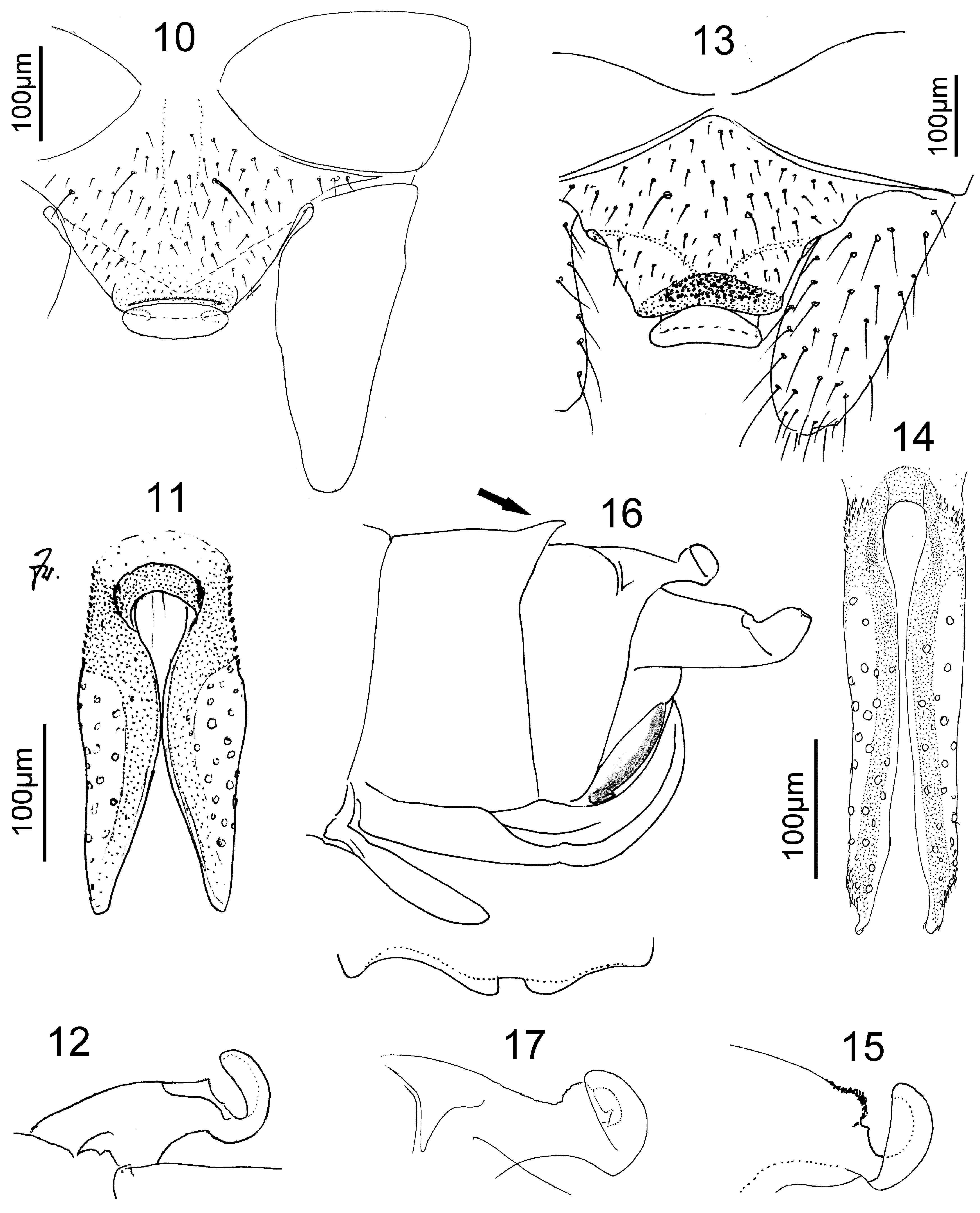
Male terminalia. Segment 9 sclerotized, pleural fold distinct. A strong basal bar (bb) supports the long and slender parallel ventral lobe (vl) with membranous ventral face. There is a membranous zone between the bar and the rest of sternite 9 (S9; e.g., Fig. 2 View FIGURES 1 – 9 ). S9 narrows at the rear segment border where it forms a long and narrow subgenital plate which curves up towards the epiproct; Paulian (1951) termed this a „ titillateur.“ The root-shaped muscular sperm pump (sp) inside the base of S9 is often visible by transparency ( Fig. 3 View FIGURES 1 – 9 ). A fine duct leads to the gonopore (gp) at the tip of the prolonged and narrow subgenital plate. Segment 10 is sclerotized dorsally and on the sides, the sternite is reduced. The antecosta is divided, the plate of tergite 10 (T10) bears caudally a baseplate supporting the epiproct. Anterolaterally the baseplate is connected to the base of the cerci. Caudally from this connection the sides of the baseplate are folded under, widen caudally, eventually meet and form a peduncle for the erect epiproct which curves forward over the edge of the baseplate, to a variable extent. In front of the epiproct is a patch of black granules on the rear part of the baseplate. In profile it is more or less raised. The epiproct itself is an erect triangular plate ( Figs. 7, 8 View FIGURES 1 – 9 ) or a wide shallow dish which is convex behind and concave in front (e.g., Fig. 15 View FIGURES 10 – 17 ). The appearance of dish-like epiprocts changes with slight differences in angle of view.
Each paraproct is divided into a large rounded lateral lobe which forms a pilose bulge medially from the cercus (not shown!) and a sclerotized median lobe which is common among Nemouroidea. The retractor muscles from S9 attach to the median lobes which can thereby be recognized even though they are separate from the outer lobes ( Fig. 4 View FIGURES 1 – 9 ). In Madenemura the median lobes have shifted far forward and lie in the intersegmental membrane 9/10, above the long subgenital plate but are separate from it. The median paraproct lobes are connected by a horseshoeshaped sclerite to which the retractor muscles from S9 attach ( Fig. 4 View FIGURES 1 – 9 ). The lobes bear circular sensilla and form the sides of an elongate paraproct body which is often visible in side view (pb, e.g., Figs. 2, 4, 9 View FIGURES 1 – 9 ). Paulian mentioned it in Tsaranemura culminalis Paulian, 1951 but believed it was the long recurved tip of the titillateur.
Female terminalia (e.g., Figs. 5, 6 View FIGURES 1 – 9 ). The tergites, cerci and paraprocts are unmodified. S8 is convex and well sclerotized, caudally it is restricted and forms a subgenital plate. S9 has a similar, slightly narrower and longer caudal extension, a postgenital plate resting on the subgenital plate. The gonopore is located at the front of the deep fold between sub- and postgenital plate which together form an ovipositor of specific length and shape.
Larva. The habitus resembles Nemouridae except that the wing pads do not diverge obliquely sideward but extend backward, parallel to the body axis. Pleural folds separate tergites from sternites except on abdominal segments 9 and 10. The legs are moderately slender, the straight hind leg extends distinctly beyond the tip of the abdomen. First tarsus segment elongate, 2nd minute, dorsally barely visible between the first and the very long 3rd segment. Antennae and cerci long and slender, no special pilosity. No gills. Mouthparts unmodified ( Paulian 1949), the wide mentum agrees with Aphanicerca ( Zwick 2006: fig. 6).
In males ( M. descarpentriesi Paulian, 1959 and M. culminalis (Paulian, 1951)) , the long subgenital plate develops in an inconspicuous bare conical pocket which projects a little beyond the segment border. It originates from the soft area dorsally from the continuous setal fringe along the rear edge of S9. In the single available pharate male of M. culminalis the dark paraproct body was clearly visible through the cuticle of S9. After the larval cuticle was removed the epiproct was found to be closely appressed to the body (similar to Fig. 17 View FIGURES 10 – 17 ), the terminal sclerite had already its definite shape.
In females, sternites 7 and 8 have a widely interrupted, S9 a continuous caudal fringe of spreading strong setae. In a mature last instar larva of M. descarpentriesi subgenital and postgenital plate of the developing adult were visible through the semitransparent sternal cuticle. The sternites were not prolonged or modified.
Specific distinction of larvae was not attempted.
Status of Tsaranemura . Originally proposed as a separate genus, later ( Paulian 1959) classified as subgenus of Madenemura with which it is here synonymized. The type and only species, M. culminalis , agrees in structure completely with the other species and is most similar to M. descarpentriesi .
Affinities. Madenemura agrees with no other genus of the family, the spindle-shaped paraproct body of males is unique. The South American fauna ( Aubert 1960, Illies 1961, Stark et al. 2009) includes no distantly similar species. Several New Zealandese ( McLellan 1991) and Australian ( Illies 1975) females, especially in Omanuperla McLellan , Austrocercella Illies and Austrocercoides Illies , also have ovipositors formed by co-apted extensions of S8 and S9. Close relations with Madenemura are improbable in view of other differences, not least the different males.
Using the key to the South African genera ( Stevens and Picker 1995) one arrives near Aphanicercopsis for both sexes. However, the detailed descriptions and figures ( Barnard 1934) show clear differences, e.g., in the male paraprocts. In the ovipositors of Aphanicercopsis and Afronemoura Illies (figures in Balinsky 1956, as Aphanicercopsis ) the long apically divided subgenital plate of S8 accomodates the much shorter postgenital plate of S9. The opposite length relation occurs in Madenemura . The female paraprocts of the two African genera are triangular, caudally much prolonged and closely appressed while those of Madenemura are short, rounded and widely separate.
Of the South African Notonemouridae larvae, tergite and sternite are separated by a pleura in most abdominal segments only in Balinskycercella Stevens & Picker (Picker & Stevens 1997) . However, larval Balinskycercella have long setae while those on the tergites of Madenemura are short. The sternites up to and including S7 are shining and only sparsely covered with minute thin hairs. However, the last segments differ, there are long stiff spreading setae ventrally and on the paraprocts. The closest relative of Madenemura is unknown.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Madenemura Paulian, 1949
| Zwick, Peter 2015 |
Tsaranemura
| Paulian 1951 |
T. culminalis
| Paulian 1951 |