
Heteralepas gettysburgensis, Lobo & Tuaty-Guerra, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.385 |
|
publication LSID |
lsid:zoobank.org:pub:47B572DF-900E-4EB4-9A78-4B8E9220FFE2 |
|
DOI |
https://doi.org/10.5281/zenodo.5689600 |
|
persistent identifier |
https://treatment.plazi.org/id/6F9E2643-558E-4799-9FA8-EBAB9B3D04B9 |
|
taxon LSID |
lsid:zoobank.org:act:6F9E2643-558E-4799-9FA8-EBAB9B3D04B9 |
|
treatment provided by |
Plazi |
|
scientific name |
Heteralepas gettysburgensis |
| status |
sp. nov. |
Heteralepas gettysburgensis sp. nov.
urn:lsid:zoobank.org:act:6F9E2643-558E-4799-9FA8-EBAB9B3D04B9
Figs 2–6 View Fig View Fig View Fig View Fig View Fig
Diagnosis
Demarcation between capitulum and peduncle clear. Capitulum wider than capitulo-peduncular junction. Capitulum slightly longer than peduncle, both with folds. Carinal margin thickened. Aperture more than ⅓ and less than ½ height of the capitulum, with crenulated lips. Lips surrounding aperture clearly set off from the surrounding capitulum by a demarcation border. Mandible with four teeth covered by fine setae, as well as its posterior side. Lower margin of the teeth with numerous pectinations: the first tooth with ten, the second, third and fourth ones with five, five and six pectinations, respectively. First maxilla with two strong acuminate teeth of unequal length on the upper angle.
Etymology
The specific epithet alludes to the location where the specimens were collected: the Gettysburg Seamount, Gorringe Bank, in the Madeira-Tore geologic complex.
Material examined
Holotype:
NORTH-EAST ATLANTIC: located approximately 200 km off the southwestern coast of Portugal, Gettysburg Seamount, Gorringe Bank, Madeira-Tore geological complex , 36º34′42.46″ N, 11º35′49.02″ W, 255 m depth, 28 Aug. 2016 ( MB11-000943 ).
GoogleMapsParatypes:
NORTH-EAST ATLANTIC: 14 specimens, same data as for holotype ( MB11-000944 – MB11-000957 ). GoogleMaps
Description
The fresh specimens were translucent yellowish, allowing observation of the cirri movement inside the capitulum. Demarcation between capitulum and peduncle clear. Capitulum wider than capitulo-
peduncular junction ( Fig. 2A–B View Fig ). Capitulum 1.2 to 1.6 times longer than peduncle, cylindrical. Both peduncle and capitulum with folds. Aperture more than ⅓ and less than ½ of the capitulum height. Lips surrounding the aperture crenulated and clearly set off from the surrounding capitulum by a demarcation border ( Fig. 2C–D View Fig ). Carinal margin thickened. Mandible ( Fig. 3A–B View Fig ) with four teeth covered by fine setae as well as its posterior side. Lower margin of the teeth with pectinations: first tooth with ten, second, third and fourth ones with five, five and six pectinations, respectively ( Fig. 3C–F View Fig ). Distance between first and second tooth 1.5 times that between second and third tooth. Latter distance similar to the distance between third and fourth tooth. First maxilla deeply notched, with two strong acuminate teeth of unequal length on upper angle, numerous setae and short to long spines on the cutting margin. Posterior margin with a dense clump of fine setae. ( Fig. 4A–C View Fig ).Second maxilla with two lobes covered with numerous long serrulate setae, those on posterior lobe longer than those on anterior lobe ( Fig. 4D–E View Fig ). Cirral setation lasiopod. Cirrus I separated from posterior pairs, with unequal rami.Anterior ramus slightly shorter and wider at base than posterior ramus ( Fig. 5A–B View Fig ). Both with numerous setae on upper and lower margins. Base of cirrus I with one filamentous appendage. Cirrus II with anterior ramus slightly shorter than posterior ramus ( Fig. 6A View Fig ). Anterior and posterior rami of cirri III and IV very similar ( Fig. 6B–C View Fig ). Cirri V and VI similar ( Fig. 6D View Fig and 4C–D View Fig , respectively), with inner rami atrophied. Inner ramus of cirrus VI shorter than inner ramus of cirrus V. Caudal appendage slightly longer than pedicel of cirrus VI ( Fig. 5E–F View Fig ). Penis relatively long, annulated, with numerous long and short fine setae distally scattered throughout ( Fig. 5G–H View Fig ). Table 1 View Table 1 displays the measurements, ratios, mean, standard deviation and coefficient of variation concerning the morphological characters mentioned.
Discussion
All specimens were collected from the same site, at a depth of 255 m in the northeastern Atlantic. Seven species occur in the Atlantic: H. belli ( Gruvel, 1902) , H. cantelli Buhl-Mortensen & Newman, 2004 , H. cornuta (Darwin, 1851) , H. lankesteri ( Gruvel, 1900) , H. luridas Zevina, 1975 , H. microstoma (Gruvel, 1901) and H. segonzaci Young, 2001 , of which only three species occur in the east Atlantic ( H. cornuta , H. microstoma and H. segonzaci ). The new species differs distinctly from H. cornuta due to the absence of conspicuous triangular projections in the capitulum ( Fig. 2A–B View Fig ). The species H. microstoma has a peduncle much longer than the capitulum, being the opposite of our specimens, where the capitulum is slightly longer than the peduncle. The original description by Gruvel (1902) gives an account of the number of articles of cirrus I (inner and outer rami), the inner rami of cirri V and VI and the caudal appendage. All cirri have a greater number of articles than our specimens except the caudal appendage ( Table 2). The peduncle of H. segonzaci , from a depth of 2235 m, is three times shorter than the capitulum and the aperture is more than half the length of the capitulum ( Young, 2001) while our specimens have an aperture length less than half that of the capitulum, and the rami of cirri III and IV have fewer articles than H. segonzaci . Comparing with the other species recorded from the western Atlantic, H. belli has the capitulum shorter than the peduncle and the cuticle is almost smooth, with some irregular folds ( Gruvel, 1902), while our specimens have the capitulum longer than the peduncle and the cuticle with numerous folds. Moreover, in H. bellii the aperture is half as high as the capitulum height ( Gruvel, 1902) while in our species it is less than half its height. In H. lankesteri the capitulum is approximately as long as the peduncle (ratio length of capitulum to length of peduncle (C/P) varies from 0.6 to 1.3) ( Gruvel, 1900), while in our specimens the ratio varies between 1.2 and 1.6. The cirri rami with a different number of articles and the non-pectinated mandible of H. lankesteri ( Gruvel, 1900) also distinguish it from our species ( Table 2). Finally, the small size of H. luridas (maximum registered size 9.5 mm), from the Caribbean, and the smaller number of articles of all cirri rami ( Zevina, 1975) strongly distinguish it from our specimens ( Table 2).
Regarding the species outside of the Atlantic, H. japonica Aurivillius, 1892 and H. canci Chan, Tsang & Shih, 2012 are the most similar species externally, probably due to the large morphological variability of H. japonica , unlike the species analyzed in this study ( Table 1 View Table 1 ). However, they have crests on the carinal region of the capitulum, the rami of the cirri have fewer articles than our specimens and the ratio ‘height of aperture/height of capitulum’ is higher ( Chan et al., 2009). The teeth of the mandible of H. japonica appear to be without pectinations or might have low pectinations on the lower margins of teeth 1–3, especially 2 and 3 ( Buhl-Mortensen & Newman 2004), while the new species exhibits numerous pectinations on the lower margin of the four teeth. Also, the maximum length of the peduncle and capitulum of H. japonica reported by Buhl-Mortensen & Newman (2004) is 11.6 and 3.6 cm respectively, quite distinct from the new species described in this study (1.4 and 1.6 cm, respectively). On the other hand, the sequences of the COI obtained for our specimens had divergences of 16 to 21 % with H. japonica and 16 to 17% with H. canci ( Chan et al. 2009) . The intraspecific distances obtained for our specimens were 0.3 to 0.7%. Many studies have been carried out using the mitochondrial DNA sequences of the COI-5P, which have confirmed that DNA barcodes ( Hebert et al. 2003) are a reliable tool to discriminate species of crustaceans ( Lobo et al. 2013, 2016), including cirripedes ( Chan et al. 2009). Ratnasingham & Hebert (2013) suggested that 2.2% of average intraspecific distance is a reference threshold for within-species boundaries, which is quite far from the divergences found in this study. In addition, the marker 12S presented divergences approximately of 6.5% and 7.5 %, with the species H. canci and H. japonica , respectively, being within the expected values for barnacle species within the same genus ( Chan et al. 2007, 2009). The marker 16S also showed 7 % of genetic divergence with the only specimen of Heteralepas (unidentified) available in public databases ( Schiffer & Herbig 2016, GenBank accession KT947465 View Materials ), which is in accordance with the results obtained for other crustaceans ( Brasher et al. 1992; Machado et al. 1993). H. adiposa Zevina, 1982 and H. cygnus Pilsbry, 1907 , are similar to H. microstoma , but they differ from it in having numerous small calcareous knobs, four mandible teeth, and a great number of caudal segments ( Zevina & Kolbasov 2000). Therefore, their diagnostic characters are also distinct from the specimens analyzed in this study. H. mystacophora Newman, 1964 has a smooth capitulum and the superior margins of the second and third teeth of the
mandible support several widely spaced spinules ( Zullo & Newman, 1964), while the species described here presents a wrinkled capitulum and has spinules in the inferior margin in the four teeth of the mandible. The capitulum of H. nicobarica Annandale, 1909 is indistinctly separated from the peduncle, the aperture height is ¼ of the capitulum height and the peduncle is similar to or longer than the capitulum ( Annandale, 1909). Contrarily, the new species presents a clear demarcation between capitulum and peduncle, the aperture height is> ⅓ and <½ of the capitulum height and the peduncle is shorter than the capitulum (see Table 1 View Table 1 ). In H. rex ( Pilsbry, 1907) , the capitulum is almost as long as the peduncle, and lips are slightly crenulated or irregularly warty and do not protrude in adults ( Pilsbry, 1907). In our specimens the capitulum is slightly longer than the peduncle and the lips are clearly crenulated and protruding. In H. utinomii Newman, 1960 , the length of the peduncle is one third of that of the capitulum ( Newman, 1960), which distinguishes it from our specimens whose peduncle is only slightly shorter than the capitulum.
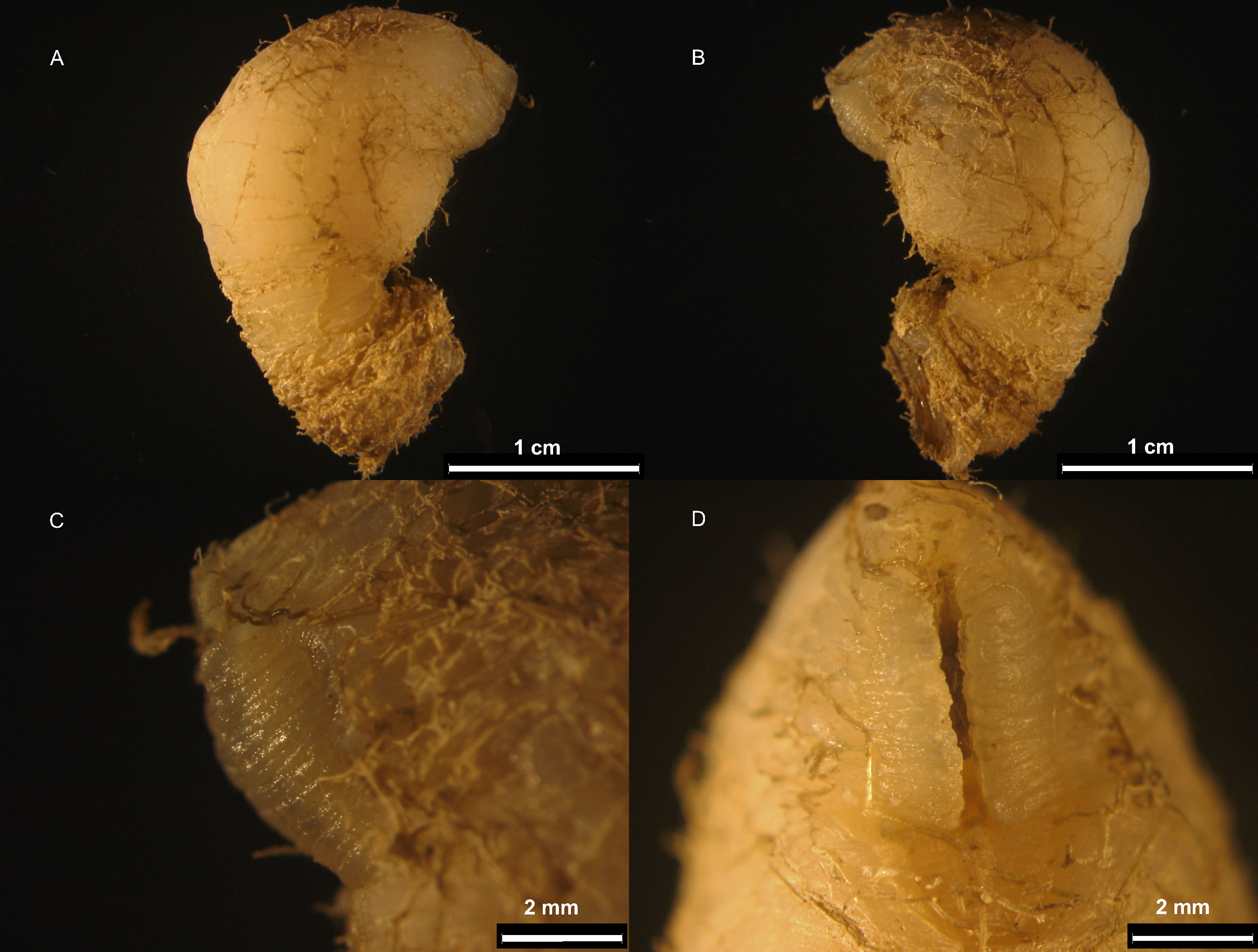

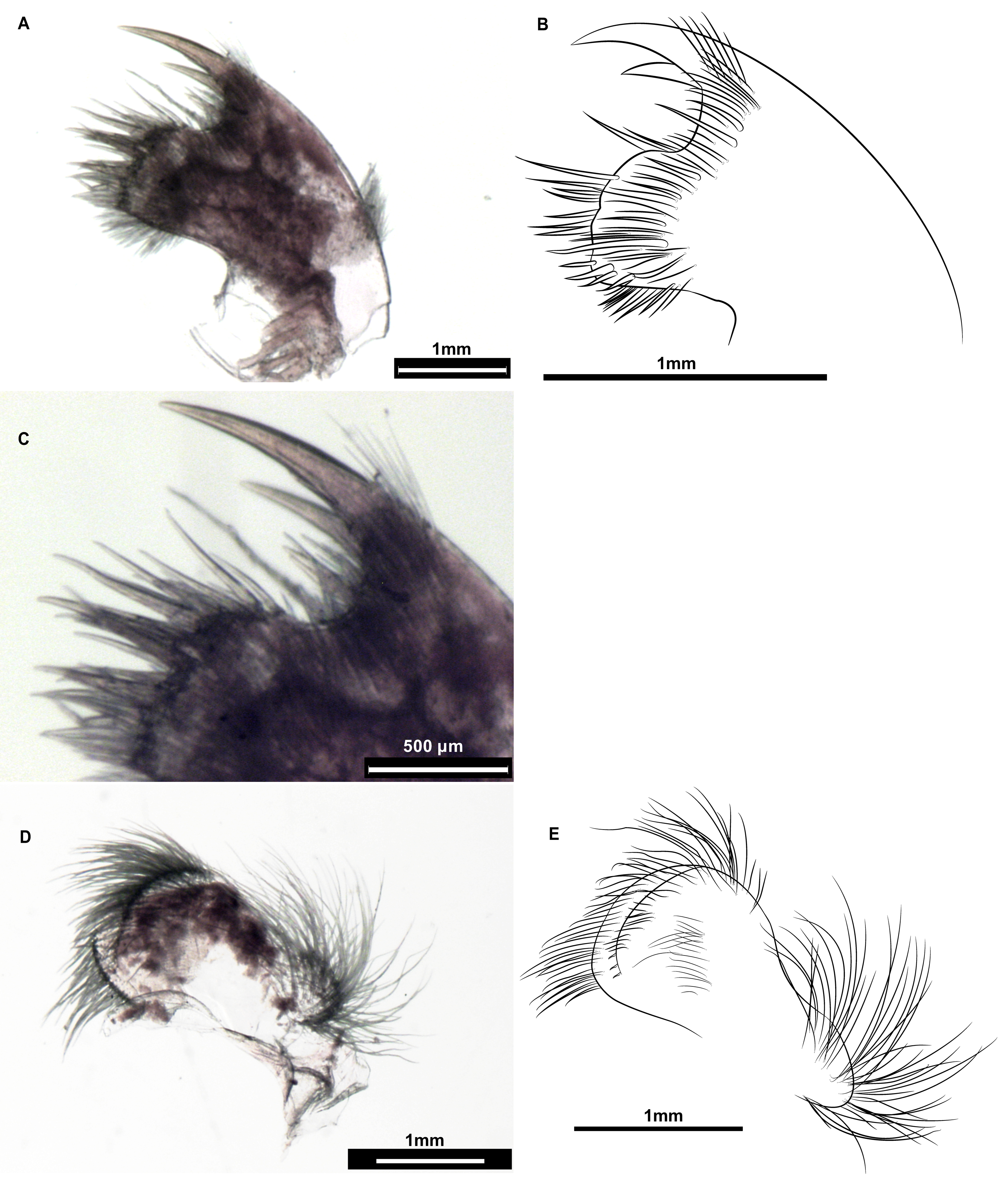

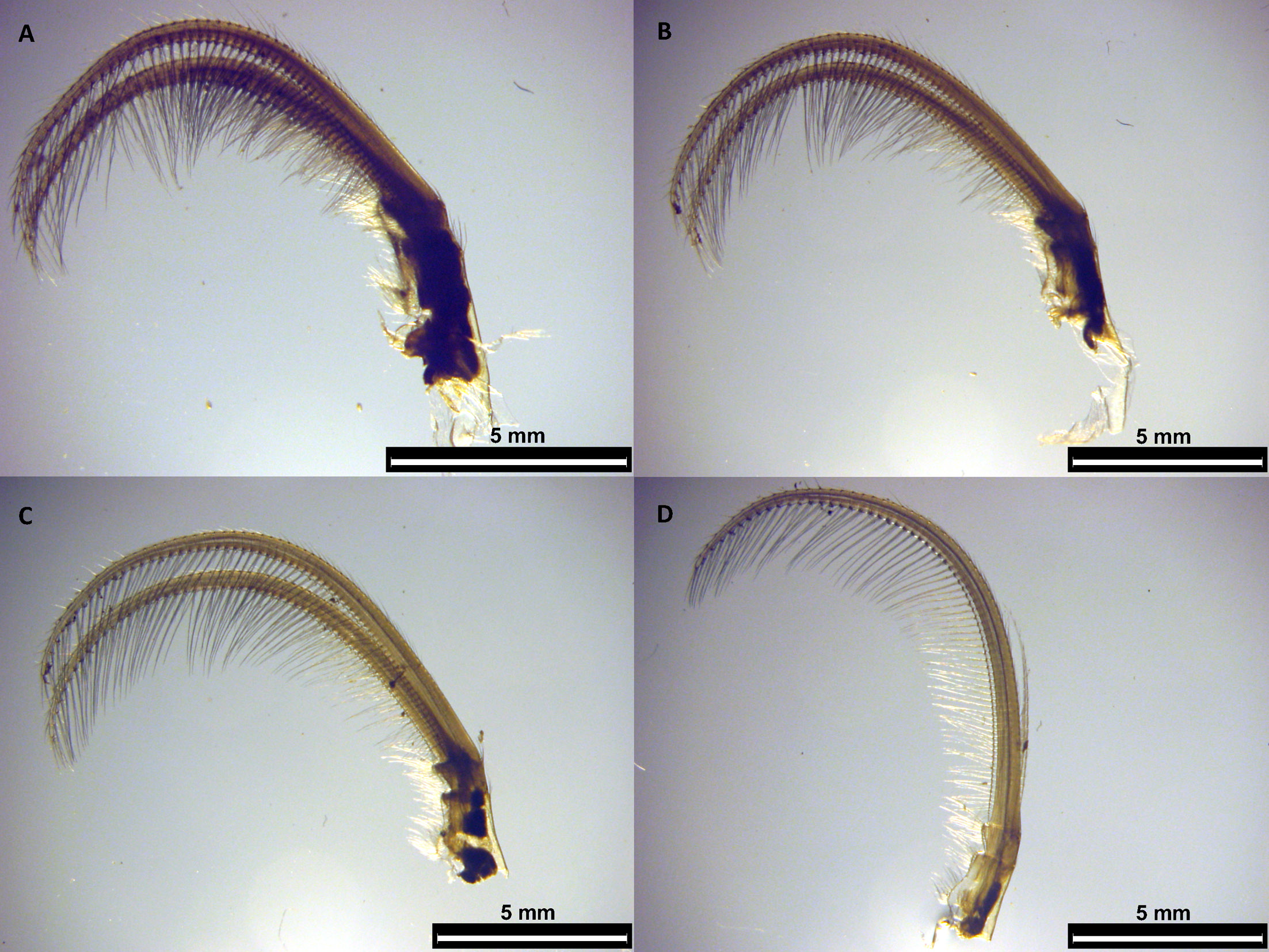
Table 1. External and internal morphological variation for specimens analyzed in this study. (- lack of information.)
| H1 | H2 | H3 | H4 | H5 | H6 | H7 | H8 | H9 | H10 | Mean | Std dev | Coefficient of variation | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Capitulum | Width | 0.87 | 1.1 | 0.88 | 0.95 | 1.08 | 0.92 | 1.12 | 0.96 | 0.96 | 1.18 | 1.0 | 0.1 | 0.09 |
| Length | 1.47 | 1.45 | 1.33 | 1.5 | 1.55 | 1.34 | 1.54 | 1.53 | 1.38 | 1.58 | 1.5 | 0.1 | 0.05 | |
| Peduncle | Width | 0.76 | 0.75 | 0.64 | 0.72 | 0.84 | 0.82 | 0.81 | 0.82 | 0.81 | 0.93 | 0.8 | 0.1 | 0.07 |
| Length | 1.1 | 0.91 | 0.97 | 1.03 | 1.28 | 1.13 | 1.08 | 1.19 | 0.95 | 1.36 | 1.1 | 0.1 | 0.10 | |
| C/P | 1.34 | 1.59 | 1.371 | 1.456 | 1.21 | 1.19 | 1.43 | 1.29 | 1.45 | 1.16 | 1.3 | 0.1 | 0.08 | |
| Aperture | Length | 0.61 | 0.47 | 0.5 | 0.57 | 0.57 | 0.49 | 0.58 | 0.54 | 0.56 | 0.69 | 0.6 | 0.0 | 0.08 |
| A/C | 0.41 | 0.32 | 0.376 | 0.38 | 0.37 | 0.37 | 0.38 | 0.35 | 0.41 | 0.44 | 0.4 | 0.0 | 0.06 | |
| Cirrus I Articles | Inner ramus | 13 | 15 | 14 | 13 | 14 | 16 | 14 | 15 | 14 | 15 | 14.3 | 0.8 | 0.05 |
| Outer ramus | 28 | 28 | 28 | 28 | 28 | 28 | 29 | 24 | 30 | 28 | 27.9 | 0.8 | 0.03 | |
| Cirrus II Articles | Inner ramus | 51 | 50 | 46 | 52 | 51 | 53 | 51 | 48 | 53 | 51 | 50.6 | 1.6 | 0.03 |
| Outer ramus | 66 | 58 | 59 | 61 | 59 | 63 | 64 | 58 | 64 | 59 | 61.1 | 2.5 | 0.04 | |
| Cirrus III Articles | Inner ramus | 65 | 67 | 61 | 65 | 69 | 67 | 66 | 62 | 68 | 61 | 65.1 | 2.3 | 0.04 |
| Outer ramus | 66 | 67 | 62 | 71 | 72 | 70 | 65 | 66 | 71 | 62 | 67.2 | 3.0 | 0.05 | |
| Cirrus IV Articles | Inner ramus | 67 | 67 | 61 | 70 | 73 | 69 | 71 | 65 | 75 | 64 | 68.2 | 3.4 | 0.05 |
| Outer ramus | 71 | 68 | 69 | 73 | 75 | 71 | 71 | 66 | 75 | 74 | 71.3 | 2.4 | 0.03 | |
| Cirrus V Articles | Inner ramus | 69 | 70 | 64 | 72 | 73 | - | 73 | 71 | 78 | 67 | 70.8 | 2.9 | 0.04 |
| Outer ramus | 22 | 19 | 17 | 22 | 27 | 19 | 26 | 21 | 19 | 20 | 21.2 | 2.4 | 0.12 | |
| Cirrus VI Articles | Inner ramus | 70 | 69 | 60 | 71 | 76 | 70 | 73 | 64 | 78 | 67 | 69.8 | 3.8 | 0.06 |
| Outer ramus | 20 | 18 | 17 | 17 | 17 | 17 | 21 | 20 | 18 | 19 | 18.4 | 1.3 | 0.07 | |
| Caudal appendage | 13 | 13 | 13 | 13 | 16 | 13 | 16 | 13 | 11 | 8 | 12.9 | 1.4 | 0.11 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |