
Trachysphaera cristangula ( Attems, 1943 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5047.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:5350CFD5-E2F1-4D42-B072-365078C11F75 |
|
persistent identifier |
https://treatment.plazi.org/id/03FBE15F-2D68-1B44-F08F-DE22FB468F0D |
|
treatment provided by |
Plazi |
|
scientific name |
Trachysphaera cristangula ( Attems, 1943 ) |
| status |
|
3. Trachysphaera cristangula ( Attems, 1943)
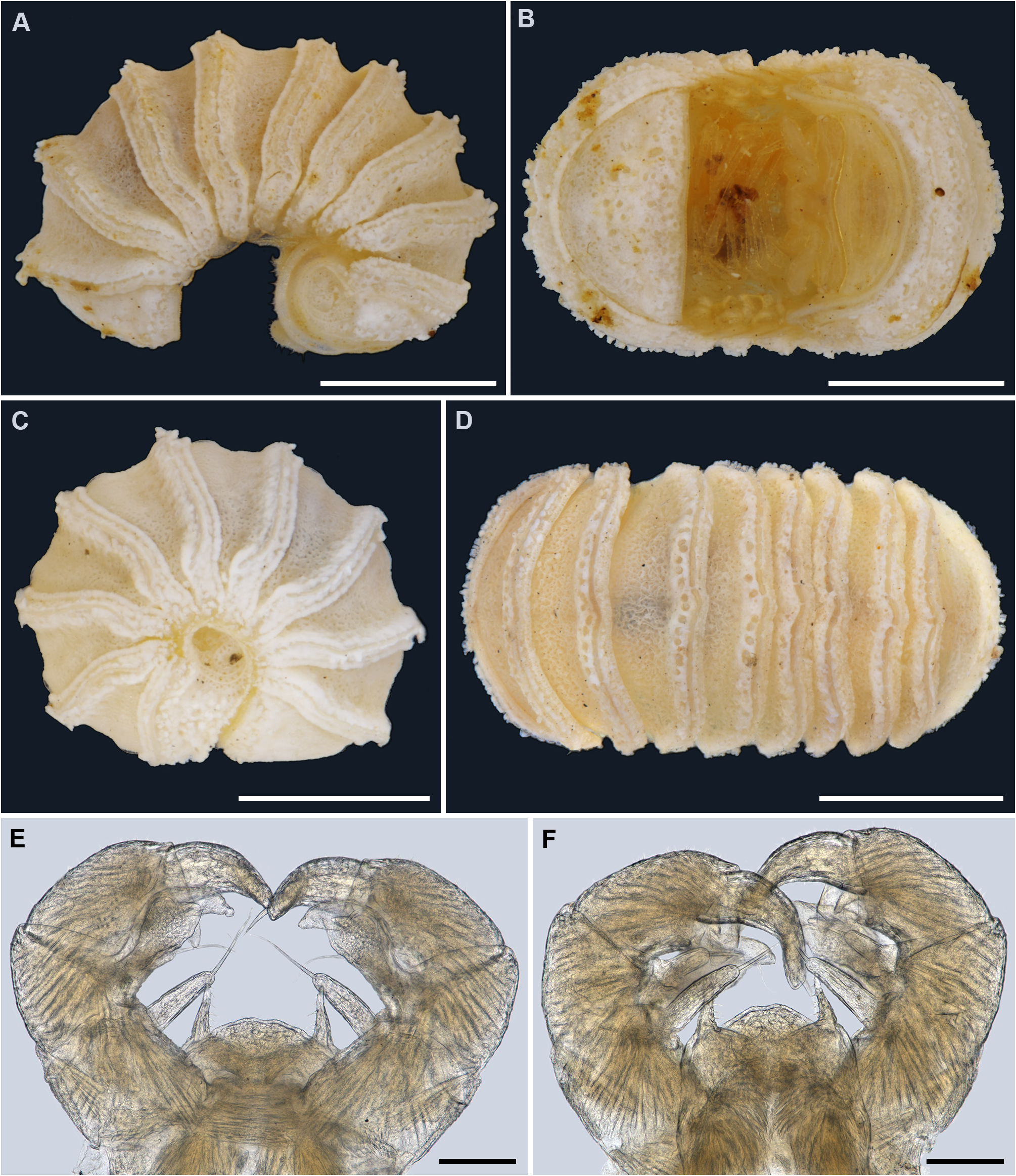
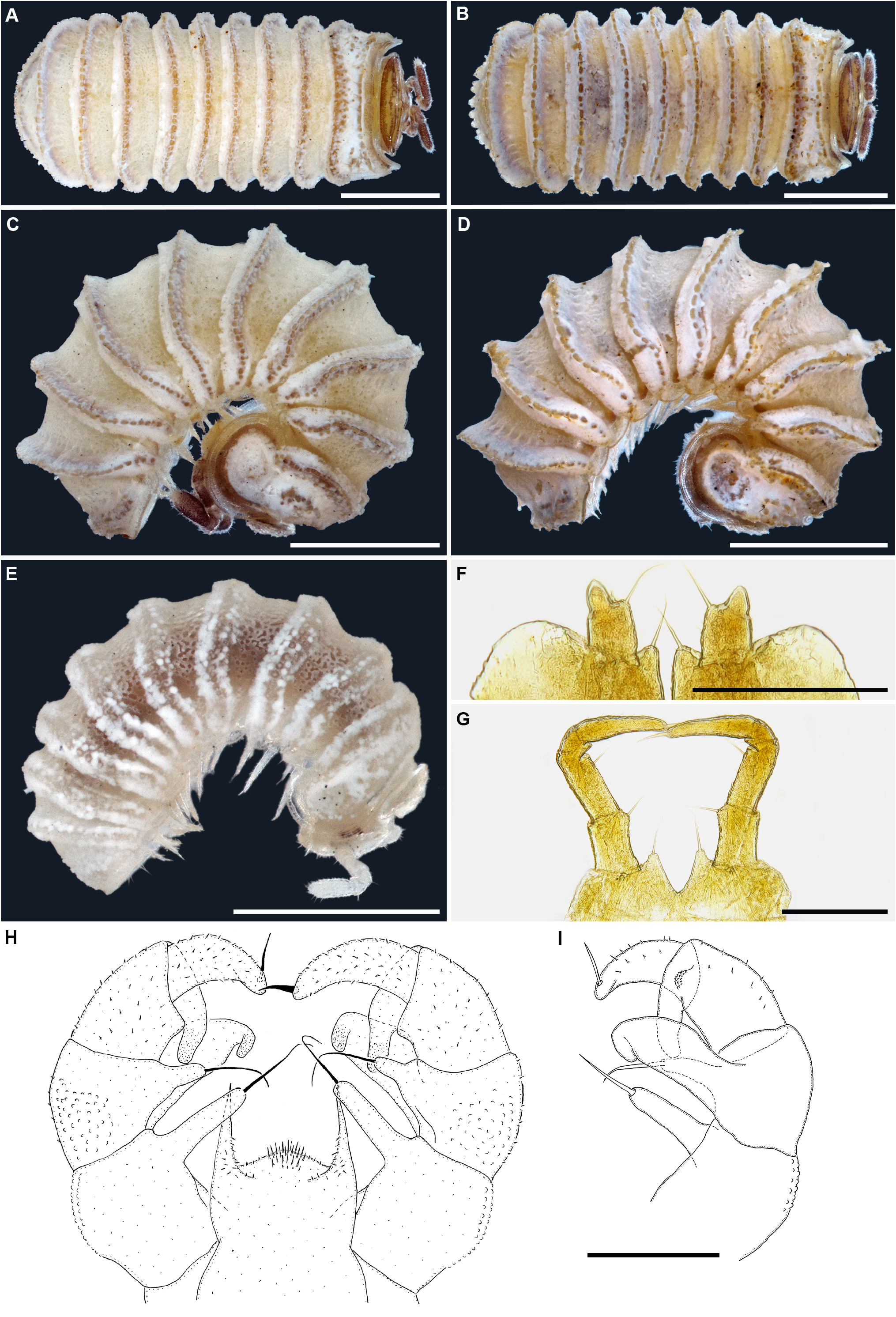
Figs 1A–C View FIGURE 1 , 5 View FIGURE 5 , 12A View FIGURE 12 .
Gervaisia cristangula Attems, 1943: 79 View in CoL , 81, 82, figs 5–8.
Gervaisia cristangula View in CoL — Attems (1929: 313), nomen nudum; Radu & Ceuca (1951: 147–149, figs 1–5).
Trachysphaera cristangula — Tabacaru (1988: 58, 59); Kime & Enghoff (2011: 36, 126).
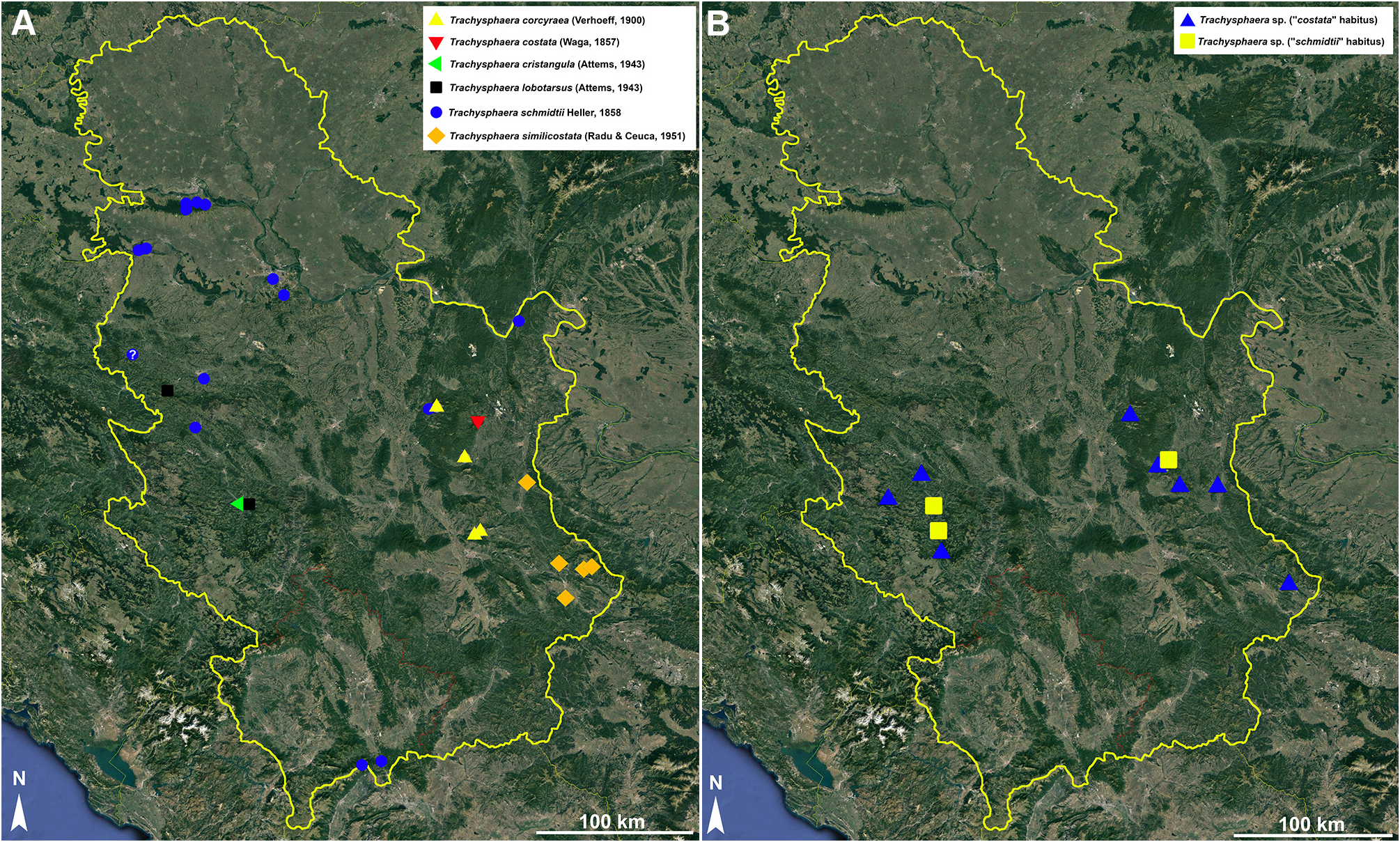
Material examined. Serbia ( IZB): 2 ♂♂, 1 ♀, near Ivanjica, 16.10.1994 ; 1 ♂, 2 ♀♀, Mucina voda, near Ivan- jica, 06.06.2021, D. Stojanović, M. Šević, D. Antić leg. Bosnia and Herzegovina ( NHMW types) : ♂ lectotype (designated herewith, NHMW MY1864 View Materials ) , 17 paralectotypes ( NHMW MY10244 View Materials ), Stolac , 1906, Penther leg. ; 34 specimens ( NHMW MY1860 View Materials , ex syntypes), Zvijezda near Vareš , 18– 20.09.1905, C. Attems leg.
Remarks. Trachysphaera cristangula had already been mentioned and recorded by Attems (1929) from several localities in Bosnia and Herzegovina, and it had remained a nomen nudum until its original description by Attems (1943: 81, 82, figs 5–8). In addition to these records, the species has also been found at a few other localities in Romania ( Radu & Ceuca 1951). The habitus ( Figs 1A–C View FIGURE 1 , 5A–D View FIGURE 5 ) and telopod structures ( Fig. 5E, F View FIGURE 5 ) of T. cristangula closely resemble those of T. schmidtii . Perhaps one of the main habitual differences is the degree to which the line of bacilliform rods/ornamentations is dorsomedially bent posteriad in T. cristangula ( Figs 1A, B View FIGURE 1 , 5D View FIGURE 5 ). This line is very obvious in adults T. cristangula , but not in juveniles, vs. remaining straight dorsomedially in T. schmidtii ( Fig. 7A, B View FIGURE 7 ); except in some rare cases where it is bent posteriad, although never so pronounced as in T. cristangula . In addition, in T. cristangula there are mainly two rows of coagulated bacilliform outgrowths, vs. mainly one row in T. schmidtii . This latter character is however highly variable thus differences should instead be checked in the structures of the telopods. The telopods are obviously more massive in T. cristangula , characterized by a much wider syncoxite, a low medial lobe covered with sparse and very short setae, basal parts of the syncoxital horns clearly rounded laterally ( Fig. 5E, F View FIGURE 5 ) (for telopod structure in T. schmidtii , see below). Based on these differences, we lean towards the opinion that T. cristangula represents a valid species.
For the Romanian population of T. cristangula, Tabacaru (1988: 59) stated that it more likely represented T. acutula syn. n. (= T. schmidtii ). Here, we would like to highlight the problem of misidentification and different interpretations of T. acutula in the sense of Latzel (1884), as well as sensu Verhoeff (1898, 1899, 1906) and Jermy (1942). Subsequent identifications of T. acutula seem to us to have been based on Verhoeff’s (1898: fig. 8) drawings alone. Strangely enough, Verhoeff (1899: 163) examined “ Gervaisia costata , acutula (non var.!)” and stated that acutula differed from costata in the structures of the telopods, comparing his drawings from 1898, fig. 8 (of acutula ) and Latzel’s (1884, fig. 42) drawings (of costata ). However, it was clear from Latzel’s (1884: 89, figs 41, 42) description of acutula that this specimen (marked by Latzel as “α”) had high posterior ridges (unlike in the “real” costata , as depicted by him in the same work, fig. 40). Thus, Verhoeff (1899) was actually comparing “his” acutula with Latzel’s (1884) “real” acutula (= schmidtii , see under T. schmidtii ), and not with the “real” costata . Later, Verhoeff (1906: fig. 16) described a subspecies, G. acutula transsylvanica Verhoeff, 1906 , that showed similar syncoxital structures as the species he ( Verhoeff 1898, 1899) had previously identified as acutula and different from Latzel’s (1884) acutula (= schmidtii ). Similar drawings of acutula in the sense of Verhoeff (1898, 1899, 1906) were presented by Jermy (1942, figs 185–189). We guess this was the reason for Tabacaru’s (1988) statement that T. cristangula from Romania was probably T. acutula sensu Verhoeff (1898, 1899 , 1906), since the drawings by Radu & Ceuca (1951: 148, figs 2, 3) really resemble Verhoeff’s ones, but not the “real” acutula . This explains the observation of Tabacaru (1988: 59) that T. cristangula could actually be Verhoeff’s acutula . After examining the material identified by Attems as acutula from Vihorlat, we can confirm that this species matches acutula in the sense of Verhoeff, but not acutula in the sense of Latzel. If this is true, then Verhoeff’s subspecific name transsylvanica has priority. To complicate the story a bit further, a doubtful species with an almost identical name, T. transylva- nica Tömösváry, 1880, is still available. Daday (1889: 73) stated that he had examined this taxon and identified it as Latzel’s (1884) acutula . This means that Tömösváry’s (1880) name T. transylvanica has priority over Latzel’s (1884) name T. acutula syn. n. (= schmidtii ). However, Jermy (1942) referred later to this species as a “ species incertae sedis ” [see also Korsós (2003)].
Even with the probability that T. cristangula and T. acutula in the sense of Verhoeff could belong to the same taxon, we cannot exclude the possibility they do not. Bearing this in mind, as well as and Tömösváry’s T. tran- sylvanica as a nomen dubium ( Korsós 2003), we opted here to assign the specimens found in western Serbia to T. cristangula , also coming closer to the type locality of the species.
This illustrates well the existing confusion, especially in the Carpathians, concerning the occurrence of T. cris- tangula/ acutula in the sense of Verhoeff and of T. acutula in the sense of Latzel (= T. schmidtii ). In order to resolve this tangle, it is necessary to revise the entire material from that region and beyond, which seems to be impossible for the time being.
In order to stabilize the species concept, a male lectotype has been designated from the type series ( NHMW MY1864 View Materials ) .
At Mucina voda, near Ivanjica, this species lives in sympatry with T. lobotarsus . Sympatry or even syntopy is known to be common in Trachysphaera species [e.g., Gruber (2009)].
The species is new to the Serbian fauna.
Type locality. Stolac , Bosnia and Herzegovina .
General occurrence. Dinarides in Bosnia and Herzegovina and Serbia, and the Carpathians. Most probably the species has a wider distribution than presently known.
Occurrence in Serbia. So far known only from near Ivanjica, western Serbia ( Fig. 12A View FIGURE 12 ).

| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trachysphaera cristangula ( Attems, 1943 )
| Antić, Dragan, Šević, Mirko, Macek, Oliver & Akkari, Nesrine 2021 |
Trachysphaera cristangula
| Kime, R. D. & Enghoff, H. 2011: 36 |
| Tabacaru, I. 1988: 58 |
Gervaisia cristangula
| Attems, C. 1943: 79 |
Gervaisia cristangula
| Radu, V. & Ceuca, T. 1951: 147 |
| Attems, C. 1929: 313 |