
Cyrtodactylus thalang, Grismer & Pawangkhanant & Bragin & Trofimets & Nazarov & Suwannapoom & Poyarkov, 2024
|
publication ID |
https://doi.org/10.11646/zootaxa.5437.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:03EAB984-A38F-4C8C-A9A3-06D8318F6353 |
|
DOI |
https://doi.org/10.5281/zenodo.10994251 |
|
persistent identifier |
https://treatment.plazi.org/id/49B6986E-BD6B-4303-9263-1183DE16A3F7 |
|
taxon LSID |
lsid:zoobank.org:act:49B6986E-BD6B-4303-9263-1183DE16A3F7 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus thalang |
| status |
sp. nov. |
Cyrtodactylus thalang sp. nov.
urn:lsid:zoobank.org:act:
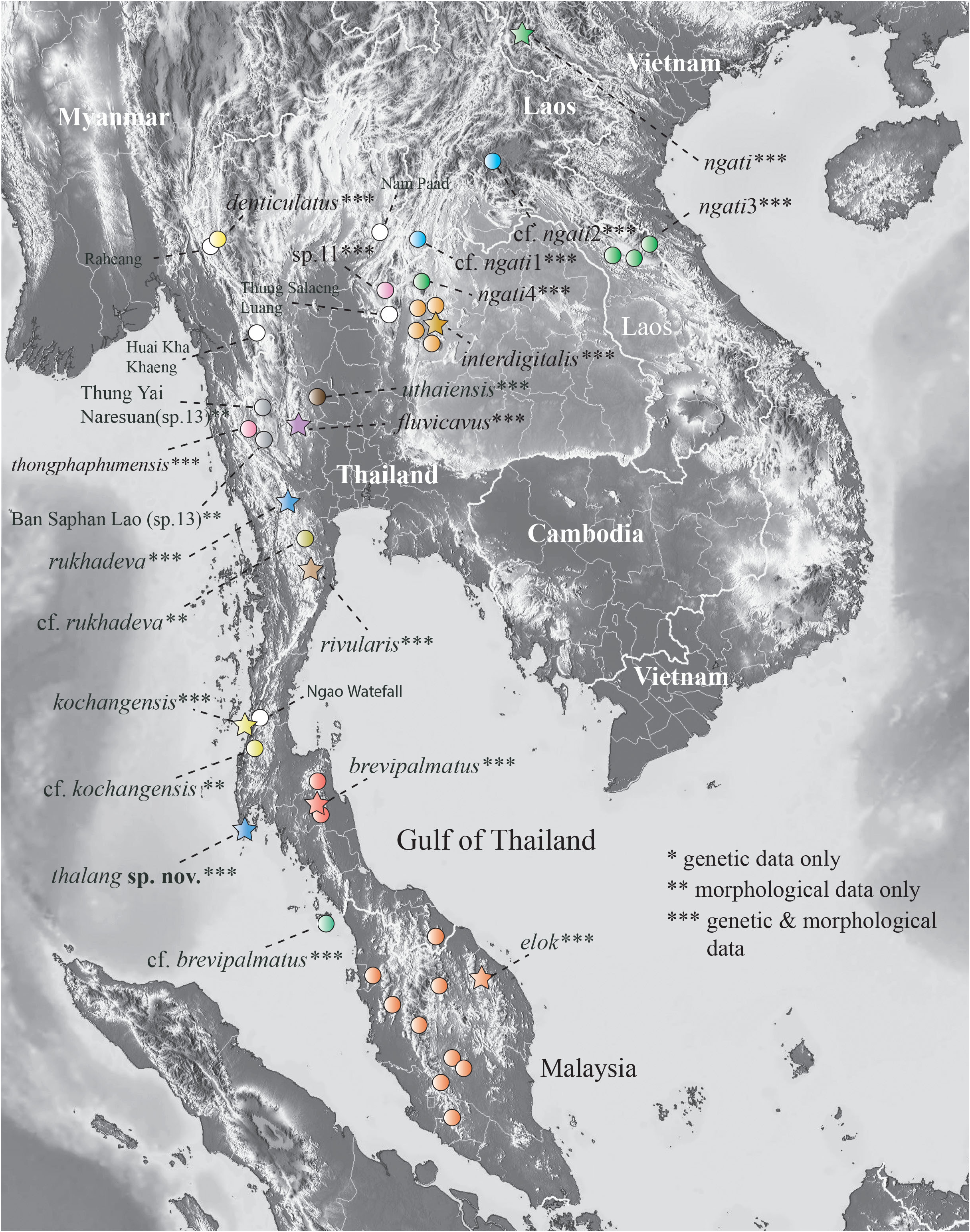
Figs. 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 ; Table 1 View TABLE 1
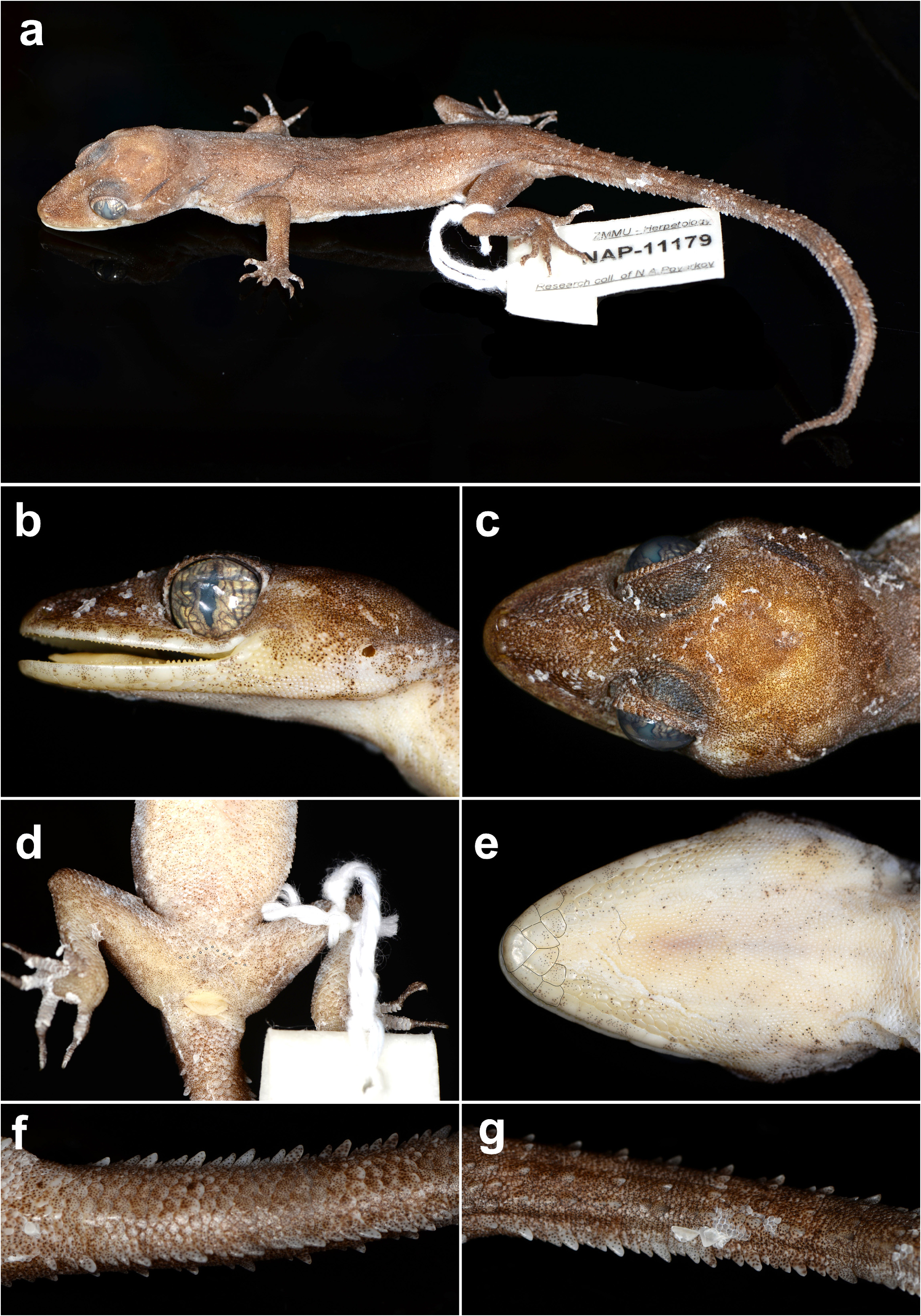
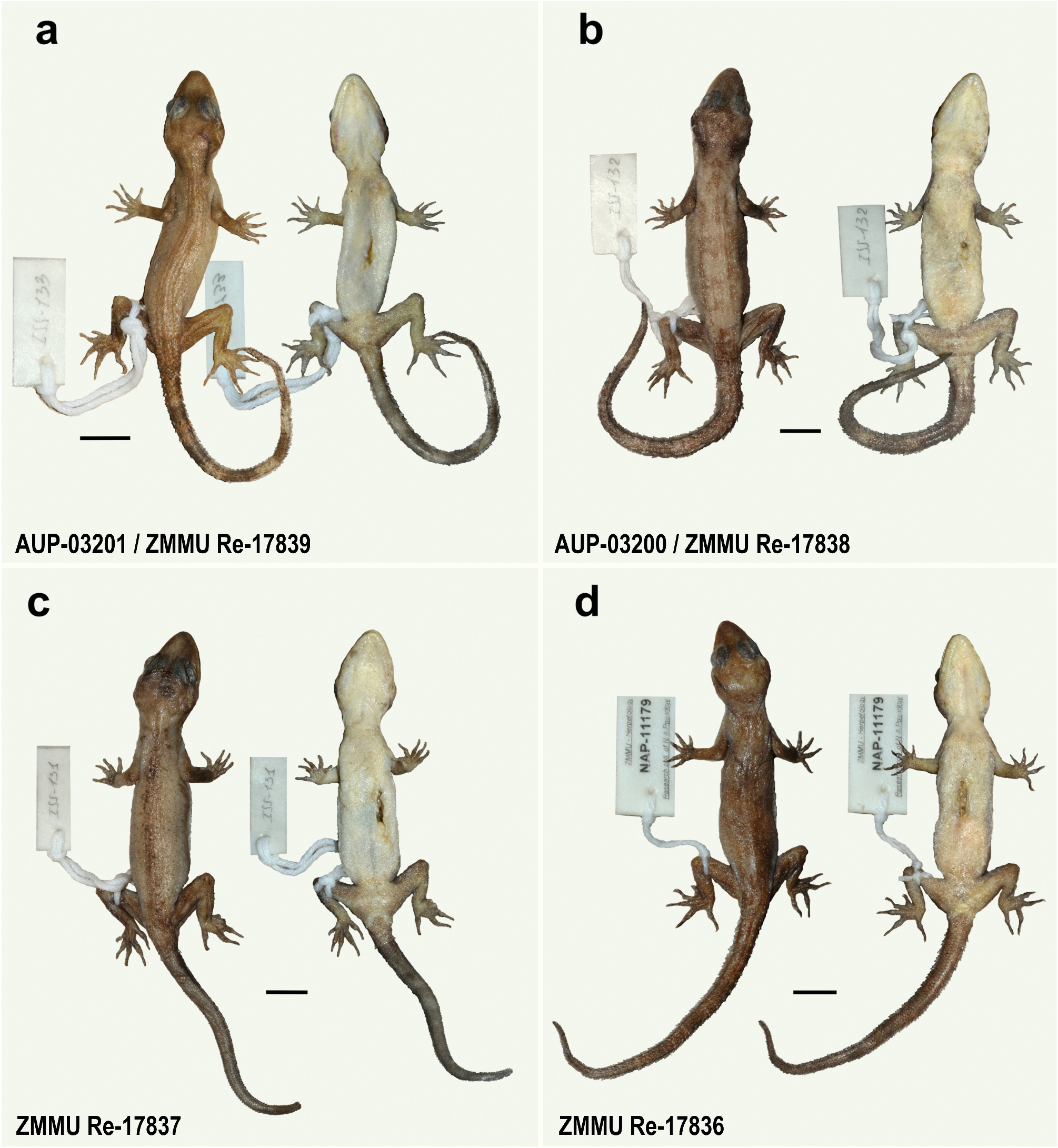
Holotype. Adult female ( ZMMU Re-17836; field tag NAP-11179) collected from Kathu waterfall , on the border of Thalang and Kathu districts, Phuket Province, Thailand ( 7.934°N, 98.323°E; elevation 112 m a.s.l.) on January 14, 2022, at 19:30 h by Poyarkov N. A., Bragin A. M., Trofimets A. V., and Suwannapoom C. GoogleMaps
Paratypes (n=3). ZMMU Re-17837 (field tag ISS-131) , ZMMU Re-17838 (ISS-132) , and ZMMU Re-17839 (field tag ISS-133), GoogleMaps three adult females collected at the same locality as the holotype on January 18, 2023, at 20:00 h by Pawangkhanant P., Suwannapoom C., Naiduangchan M., and Idiiatullina S. S GoogleMaps .
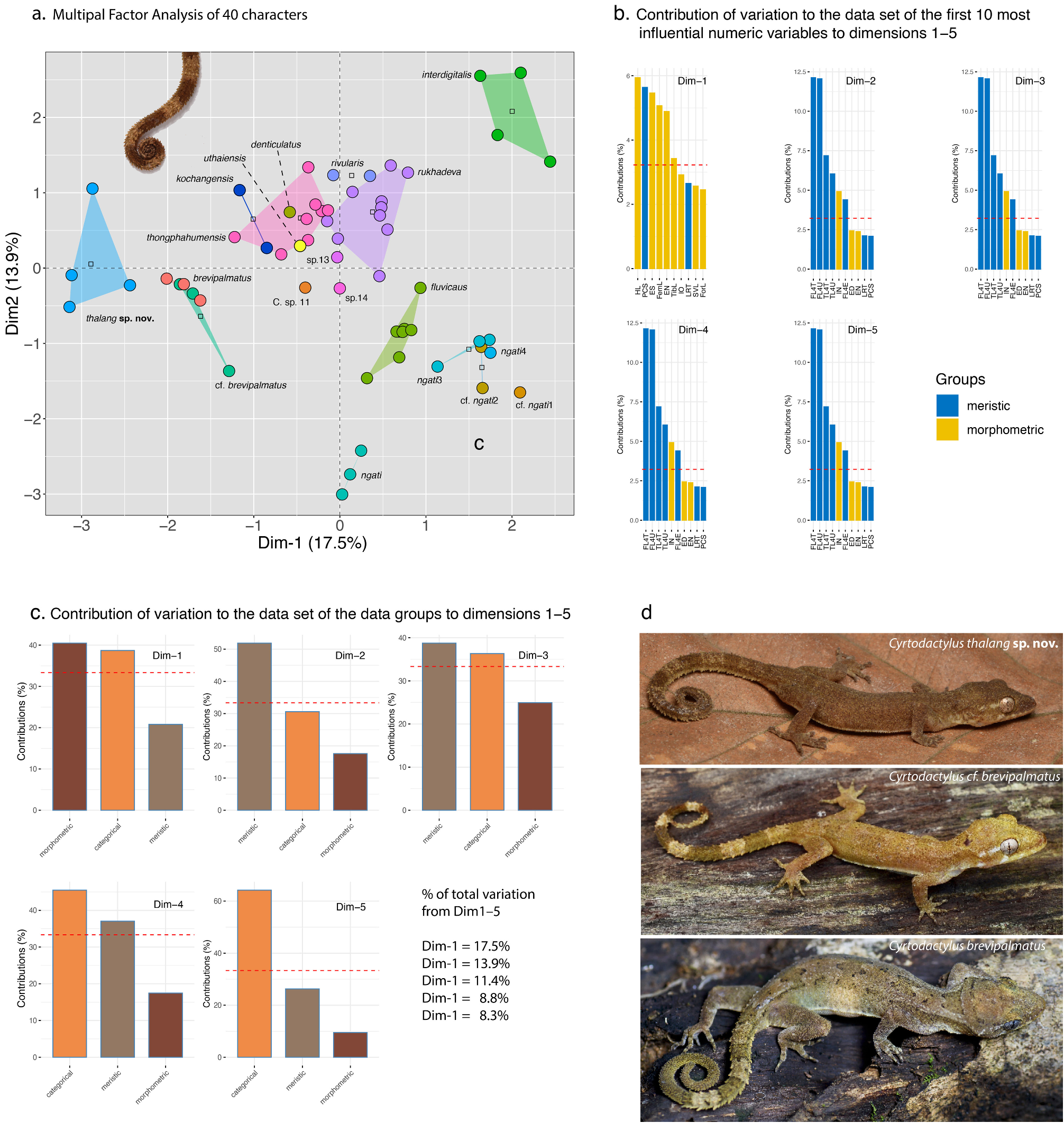
Diagnosis (females only). Cyrtodactylus thalang sp. nov. can be separated from all other species of the brevipalmatus group by the combination of having 10–12 supralabials, 9–11 infralabials, 24–31 paravertebral tubercles, 16 or 17 rows of longitudinally arranged tubercles, 28–34 longitudinal rows of ventrals, 160–169 transverse rows of ventrals, 9–12 expanded subdigital lamellae on the fourth toe, 8–11 unexpanded subdigital lamellae on the fourth toe, 17–23 total subdigital lamellae on the fourth toe; six or seven expanded subdigital lamellae on the fourth finger, 10–12 unexpanded subdigital lamellae on the fourth finger, 17 or 18 total subdigital lamellae on the fourth finger; 13–15 total number of enlarged femoral scales, 6–9 enlarged precloacals; enlarged femorals and enlarged precloacals not continuous; proximal femorals smaller than distal femorals; no tubercles on forelimbs; small tubercles on flanks; ventrolateral body fold not denticulate; large dorsolateral caudal tubercles; wide ventrolateral caudal fringe; ventrolateral caudal fringe composed scales of different size; tail square in cross-section; no ventrolateral subcaudal ridge; slightly enlarged unpaired medial subcaudals not posteromedially furrowed; maximum SVL 70.3 mm; 0–3 dark transverse body bands ( Table 1 View TABLE 1 ).
Description of holotype ( Fig. 5 View FIGURE 5 ; Table 1 View TABLE 1 ). Adult female SVL 64.5 mm; head moderate in length (HL/SVL 0.29), width (HW/HL 0.68), depth (HD/HL 0.37), distinct from neck, triangular in dorsal profile; lores slightly concave anteriorly, weakly inflated posteriorly; prefrontal region concave; canthus rostralis rounded; snout elongate (ES/HL 0.38), rounded in dorsal profile; eye large (ED/HL 0.26); ear opening oval, small; eye to ear distance greater than diameter of eye; rostral rectangular, bordered posteriorly by large left and right supranasals and three small azygous internasals, bordered laterally by first supralabials; external nares bordered anteriorly by rostral, dorsally by large supranasal, posteriorly by two unequally sized smaller postnasals, bordered ventrally by first supralabial; 10R/10L rectangular supralabials, second through sixth supralabials slightly larger than first, then tapering below eye; 10R/10L infralabials tapering smoothly to just below and slightly past posterior margin of eye; scales of rostrum and lores flat to slightly domed, larger than granular scales on top of head and occiput; scales of occiput intermixed with distinct, small tubercles; superciliaries subrectangular, largest anterodorsally; mental triangular, bordered laterally by first infralabials and posteriorly by large left and right trapezoidal postmentals contacting medially for ~85% of their length posterior to mental; single enlarged, square to rectangular sublabial; much smaller sublabials extending posteriorly to fourth(L) and fifth(R) infralabial; gular and throat scales small, granular, grading posteriorly into slightly larger, flatter, smooth, imbricate, pectoral and ventral scales.
Body relatively short (AG/SVL 0.46) with well-defined ventrolateral folds; dorsal scales small, granular interspersed with larger, weakly keeled, trihedral, semi-regularly arranged, tubercles; tubercles extend from occipital region to slightly beyond base of tail transforming in to large, nearly spine-like tubercles in paravertebral and lateral rows; smaller tubercles extend anteriorly onto nape and occiput, diminishing in size anteriorly; approximately 16 longitudinal rows of tubercles at midbody; approximately 24 paravertebral tubercles; tubercles on flanks; 28 longitudinal rows of flat, imbricate, ventral scales much larger than dorsal scales; 165 transverse rows of ventral scales; 11 large, pitted, precloacal scales; no deep precloacal groove or depression; and two rows of enlarged postprecloacal scales on midline.
Forelimbs moderate in stature, relatively short (ForL/SVL 0.15); granular scales of forearm slightly larger than those on body, not interspersed with tubercles; palmar scales rounded, slightly raised; digits well-developed, relatively short, inflected at basal interphalangeal joints; digits narrower distal to inflections; subdigital lamellae wide, transversely expanded proximal to joint inflections, narrower transverse lamellae distal to joint inflections; claws well-developed, claw base sheathed by a dorsal and ventral scale; 12R/12L expanded and 11R/11L unexpanded lamellae beneath the fourth finger; hind limbs larger and thicker than forelimbs, moderate in length (TibL/SVL 0.16), covered dorsally by granular scales interspersed with small tubercles dorsally and posteriorly and anteriorly by flat, slightly larger, subimbricate scales; ventral scales of thigh flat, imbricate, larger than dorsals; subtibial scales flat, imbricate; one row of 6R/7L enlarged pitted femoral scales not continuous with enlarged pitted precloacal scales, terminating distally at knee; 6R/7L enlarged femoral scales; proximal femoral scales smaller than distal femorals, the former forming an abrupt union with much smaller, rounded, ventral scales of posteroventral margin of thigh; plantar scales flat, subimbricate; digits relatively short, well-developed, inflected at basal interphalangeal joints; 6R/6L wide, transversely expanded subdigital lamellae on fourth toe proximal to joint inflection extending onto sole, and 12R/12L unexpanded lamellae beneath the fourth toe distal to joint inflection; and claws well-developed, claw base sheathed by a dorsal and ventral scale.
Tail original, 78.9 mm long (TL/SVL 1.22), 5.0 mm in width at base, tapering to a point; nearly square in cross-section; dorsal scales flat, intermixed with tall, conical tubercles forming paravertebral and dorsolateral rows; large, posteriorly directed, semi-spinose tubercles forming a wide ventrolateral caudal fringe; larger scales of ventrolateral fringe occur at regular intervals; medial subcaudals small unpaired, an enlarged single medial subcaudal longitudinal row absent; no subcaudal ventrolateral folds; subcaudals, larger than dorsal caudals; base of tail lacking hemipenial swellings; 3R/3L small conical postcloacal tubercles at base of tail; and postcloacal scales flat, imbricate.
Coloration in life ( Fig. 4 View FIGURE 4 ). Dorsal ground color of head, body, and limbs nearly unicolor tan-gold; faint butterfly marking on nape; light-brown, broken, diffuse postorbital stripe between eye and ear opening; very faint light-brown speckling on digits; four wide light-brown caudal bands not encircling tail extend from base to two-thirds way down tail; ventral surfaces of body and limbs beige, generally immaculate except for faint stippling, subcaudal region generally darker; iris light-gold in color bearing dark-brown vermiculations.
Variation ( Fig. 6 View FIGURE 6 ; Table 1 View TABLE 1 ). Individuals of the type series are very similar in overall coloration and pattern. ZMMU Re-17837–38 have three faint darker dorsal bands and ZMMU Re-17837 has a fully regenerated tail. Morphometric and meristic differences within and among all species of the brevipalmatus group are listed in Table 1 View TABLE 1 .
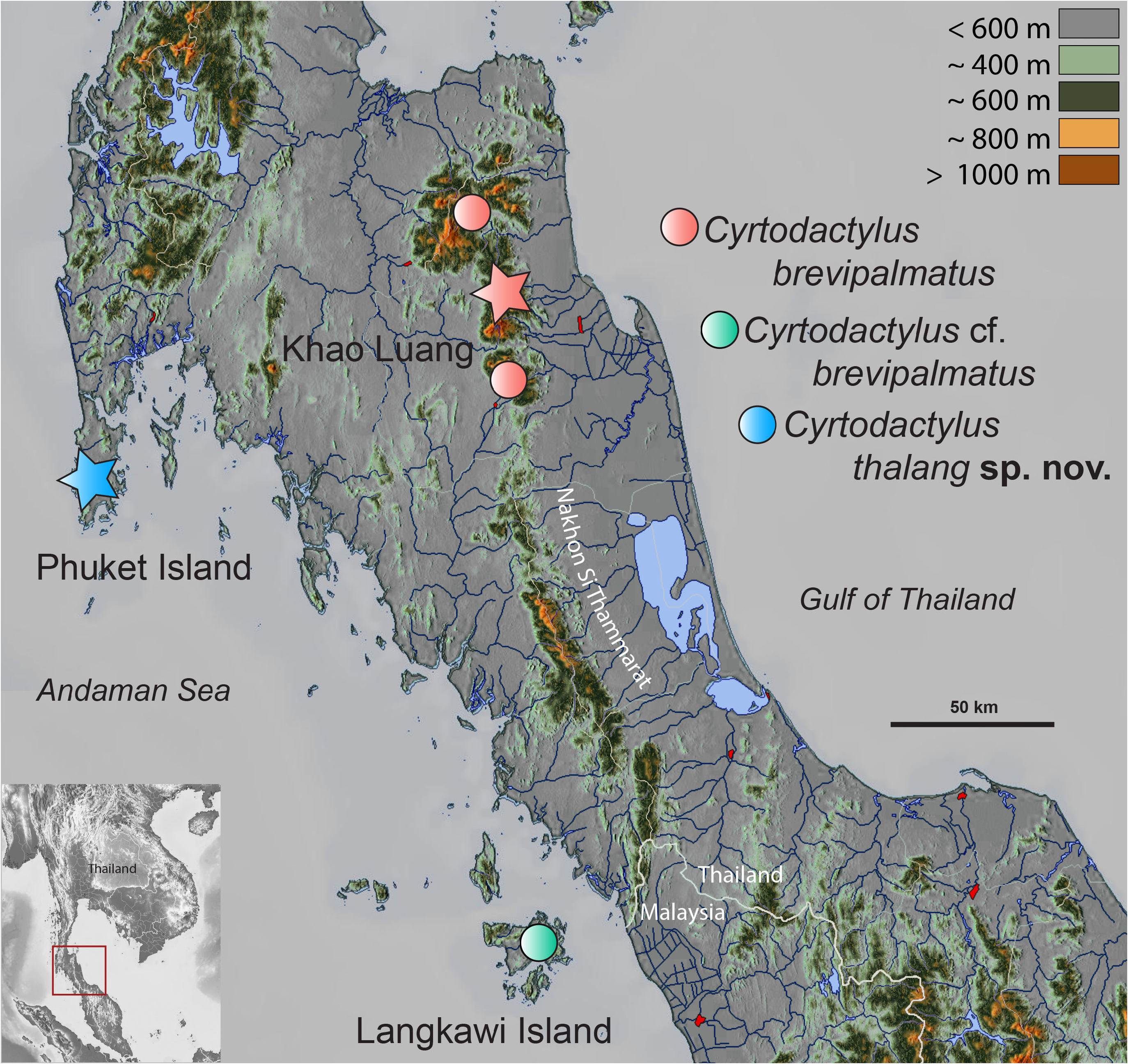
Distribution: Cyrtodactylus thalang sp. nov. is currently known only from the type locality at the Kathu waterfall, on the border of Thalang and Kathu districts, Phuket Province, Thailand in relatively close proximity to the type locality of C. brevipalmatus ( Fig. 7 View FIGURE 7 ). The fragmented upland landscape of the latter, leaves the potential for additional species (see Grismer et al. 2023a).
Etymology. The name of the new species is a noun in apposition and is therefore invariable; the species name “ thalang ” is given in reference to the historical name of Phuket Island, formerly known as Thalang (in Thai: “ถลาง”, Tha-Laang). The name also refers to the northern district of the Phuket Province, where the type locality is located. We recommend the names “ Thalang Bent-toed Gecko ” and “ Tuk-Kai Tha-Laang ” (ตุ๊กกายถลาง) as common names of the new species in English and Thai, respectively.
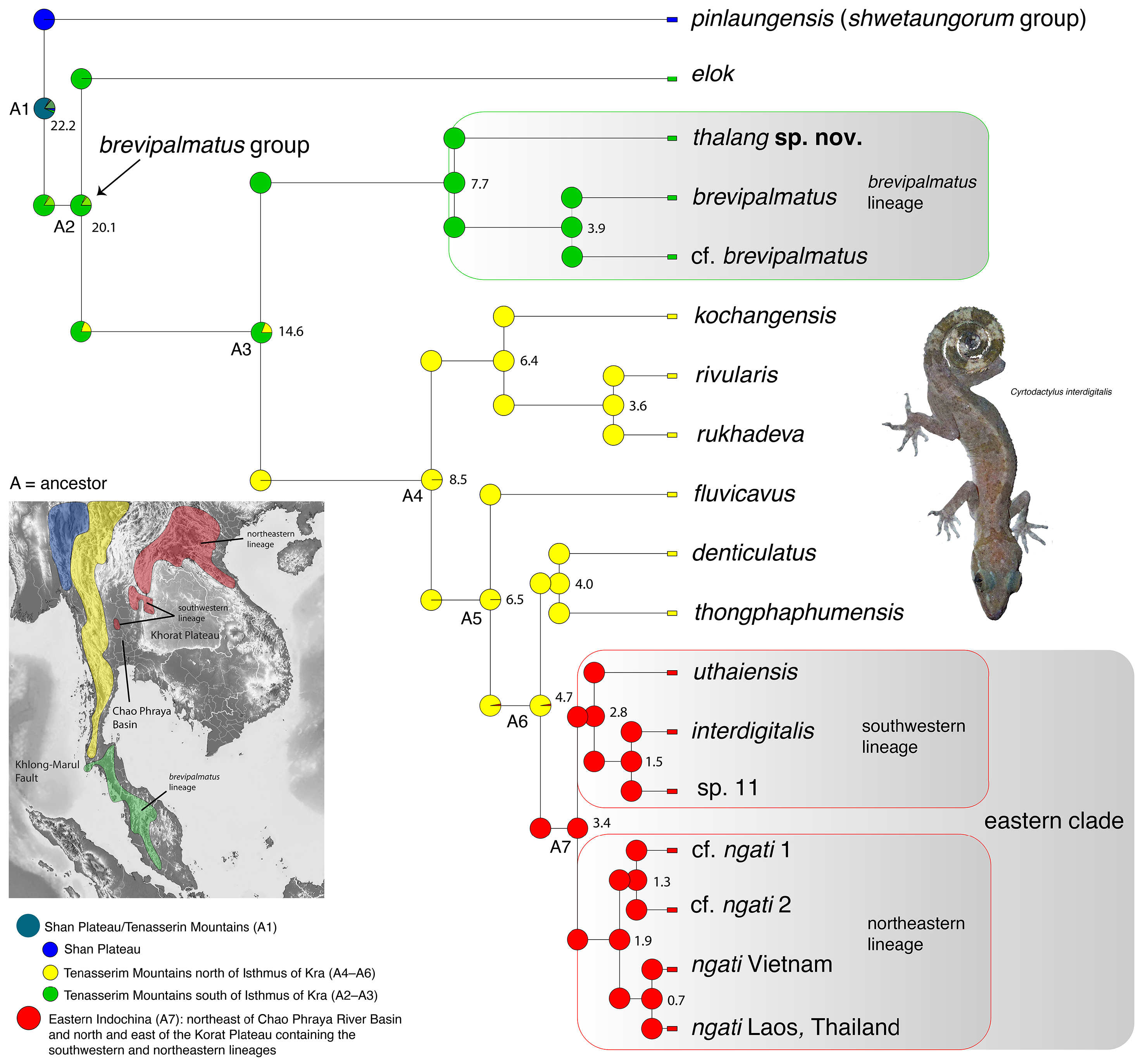
Comparisons. Cyrtodactylus thalang sp. nov. is the sister species to a lineage composed of C. brevipalmatus and C. cf. brevipalmatus ( Fig. 3 View FIGURE 3 ) and bears an uncorrected sequence divergence from them of 15.1% and 14.7%, respectively. Morphometrically, C. thalang sp. nov. differs from C. brevipalmatus and C. cf. brevipalmatus by having significantly shorter femora (FemL) and a narrower interorbital distance (IO) and differs further from C. cf. brevipalmatus by having significantly longer brachia (HumL). Meristically, C. thalang sp. nov. differs from C. brevipalmatus and C. cf. brevipalmatus by having significantly fewer ventral scales (VS), expanded subdigital lamellae on the fourth finger (FL4E), and unexpanded subdigital lamellae on the fourth toe (TL4U). It differs further from C. cf. brevipalmatus by having significantly fewer enlarged femoral scales (FS) but significantly more than in C. brevipalmatus . Categorically, C. thalang sp. nov. differs from C. brevipalmatus and C. cf. brevipalmatus by lacking as opposed to having small tubercles on the flanks (FKT), having large as opposed to small dorsolateral tubercles (DCT), a wide as opposed to a narrow ventrolateral caudal fringe (VLF1), more square as opposed to circular cross-section of the tail (TLcross), and lacking as opposed to having enlarged median subcaudals (SC2). Raw morphometric, meristic, and categorical differences among and between all other species are listed in Table 1 View TABLE 1 .
Natural history. Cyrtodactylus thalang sp. nov. inhabits lowland evergreen tropical forests, and was most often recorded in mixed bamboo and dipterocarp forests at elevations ranging from 80–200 m a.s.l. The forest where the new species was observed is dominated by the large trees of Artocarpus elasticus Reinw. ex Blume. and Dipterocarpus baudii Korth. ( Dipterocarpaceae ) ( Fig. 8 View FIGURE 8 ). Lizards were most often observed at night between 1900 h and 2400 h perched on the thin branches in the middle to upper canopy ( 6–10 m above the ground) above rocky stream. One specimen was observed ca. 2 m above the ground on a large granite boulder within a dense tangle of bamboo. At the type locality, C. thalang sp. nov. was recorded in sympatry with Cnemaspis phuketensis Das & Leong, 2004 , Cyrtodactylus cf. oldhami (Theobald, 1876) , C. macrotuberculatus Grismer & Ahmad, 2008 ( Gekkonidae ), and Tropidolaemus wagleri (Boie, 1827) ( Viperidae ).
Biogeography. Reconstructing the biogeographic history of any lineage is always problematic in the sense that missing taxa—living or extinct—can limit the accuracy of nodal probability estimates because the phylogenetic placement or the geographic location of any newly added taxon or individual can potentially overturn an entire scenario (see discussion in Grismer et al. 2023d). This potential instability is even more likely in the brevipalmatus group given that there are unsampled lineages of unknown propinquity ( Fig. 1 View FIGURE 1 ) and the likelihood of finding new, isolated upland populations throughout the mountainous regions of Indochina is all but certain. Nonetheless, below is presented the most parsimonious biogeographic scenario based on the data at hand that will form the foundation for subsequent scenarios.
The BioGeoBEARS model comparisons recovered the BAYAREALIKE model as the best fit to the data and most likely to infer the best estimates of the ancestral range at each node being that it had the lowest AIC-wt score ( Table 3 View TABLE 3 ). It is noteworthy, however, that all the trees generated from the other models, generally recovered the same ancestral range at each node, thus converging on the same biogeographical scenario, just with different levels of probability. Overall, the BAYAREALIKE model indicates that speciation within the brevipalmatus group likely progressed as a series of chronological vicariant events from south to north along the Tenasserim Mountains followed by more northerly vicariant events progressing from west to east across northern Indochina ( Fig. 2 View FIGURE 2 ). The ancestor of the shwetaungorum and brevipalmatus groups (A1) originated somewhere in the contiguous regions of the Shan Plateau and Tenasserim Mountains prior to 22.2 mybp. At approximately 22.2 mybp, A1 diverged into the shwetaungorum group which remained on the Shan Plateau in eastern Myanmar and the brevipalmatus group which spread south along the growing uplands of the Tenasserim Mountains ( Upton et al. 1995, 1997) along what is today the Thai-Malay Peninsula and thence east, through the hilly areas of northern Indochina. By ~ 20.1 mybp, the ancestor of the brevipalmatus group (A2) diverged into C. elok in the southern portion of the Tenasserim Mountains and the ancestor (A3) of the remaining species. At 14.6 mybp, additional speciation events in the southern section of the Tenasserim Mountains along the Thai-Malay Peninsula resulted in the divergence of A3 into the brevipalmatus lineage south of the Khlong-Marul Fault and the ancestor (A4) of the remaining species in Indochina. It is tempting to invoke the formation of the Isthmus of Kra region and one of its many inundations near the Khlong-Marul Fault to account for this cladogenic event given it has been responsible for divergences in this region for a vast taxonomic array of lineages (e.g. Hughes et al. 2003; de Bruyn et al. 2004; Bernstein et al. 2023). However, during this time period (Middle Miocene) there was no isthmus, and the entire western section of the Sunda Shelf was exposed ( Hall 2013). A4 began diverging in a series of chronological events along the Tenasserim Mountains from south to north across the Miocene/Pliocene border from approximately 8.5–4.7 mybp giving rise to a southern clade composed of C. kochangensis and the sister species C. rukhadeva and C. rivularis at approximately 6.4 mybp. This was followed sequentially by evolution of C. fluvicavus and the sister species C. denticulatus and C. thongphaphumensis , followed by ancestor A7 at approximately 4.7 mybp—the latter forming an eastern clade containing five or six species. This series of divergent events from the Middle Miocene onward may be the result of continued orogeny along the Thai-Malay Peninsula coupled with weak but cyclical climatic fluctuations ( Lohman et al. 2011), creating cooler upland refugia where most of these species are now restricted. Currently, the eastern clade ranges across central and eastern Indochina, northeast of the Chao Phraya Basin and north and east of the Khorat Plateau—the exception being C. uthaiensis which is restricted to an isolated karst hill in the lower Chao Phraya Basin ( Grismer et al. 2023b). The eastern clade comprises monophyletic southwestern and northeastern lineages. The presence of C. uthaiensis in the Chao Phraya Basin suggests that the eastern clade likely ranged across the basin prior to its current physiography. The divergence of A6 at approximately 4.7 mybp, coincides with of the basin’s Early Pliocene formation from an increase in riverine deltaic sediments in the north ( Choowong 2011) and marine sedimentation in the south ( Choowong et al. 2004)—ultimately morphing into a wide area of uninhabitable terrain for the particular ecological requirements of these specialized arboreal upland species. North of Chao Phraya Basin in Thailand, the eastern clade is separated from the remaining species of the brevipalmatus group by four major river systems: the Ping, Wang, Nom, and Nan —all of which drain into the upper reaches of the Chao Phraya Basin and converge in its central section in Nakkon Sawan Province. At approximately 3.4 mybp, the ancestor of the eastern clade (A7) diverged into the southwestern and northeastern lineages. These lineages are narrowly separated along the northern portion of the Pa Sak River Valley in Phetchabun Mountains of Loei Province in northern Thailand. Divergence within these lineages was recent and likely commensurate with cyclical glacioeustatic temperature changes, forcing the retreat and subsequent isolation of these species to cooler mesic upland refugia which has been reported for a number of species across the sky island archipelagos of Indochina ( Woodruff 2010; Hughes et al. 2003; de Bruyn et al. 2004; Bernstein et al. 2023).


| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |