
Periclimenaeus amirantei, Bruce, 2007
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1644.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:47F468A3-77C5-4515-A3CF-A740DC887E78 |
|
DOI |
https://doi.org/10.5281/zenodo.5103759 |
|
persistent identifier |
https://treatment.plazi.org/id/03F28784-BE52-BF15-FF0A-944A36A5CD30 |
|
treatment provided by |
Felipe |
|
scientific name |
Periclimenaeus amirantei |
| status |
|
Periclimenaeus amirantei may be distinguished from these species by the following key:
1. Second pereiopod with carpus distinctly longer than palm length, ambulatory dactyl with unguis only slightly longer than accessory tooth, R.1+7-8/1–2(3) .................................................... P. speciosus Okuno
- Second pereiopod with carpus shorter or not markedly longer than palm length, ambulatory dactyls with unguis markedly longer than accessory tooth...............................................................................................2
2. Second periopod with carpus subequal to palm length, fingers without proximal diastema, dentition 1–2/ 3–4 .................................................................................................................................................................. ................................................................................................................................... P. amirantei sp. nov.
- Second periopod with carpus shorter than palm length, both fingers with proximal diastema, dentition 1/1, R.1+7–8/2 ........................................................................................................................ P. holthuisi Bruce View in CoL
Measurements (mms). Ovigerous female holotype, postorbital carapace length, 3.9; carapace and rostrum, 7.0; total body length (approx.), 24.5; second pereiopod chela, 3.9; length of ovum, 0.5.
Host and Colouration. Unknown.
Distribution. Known only from the Amirante Islands, Seychelle Islands and North Malé Atoll, Maldive Islands.
Etymology. Named after the type locality, the Amirante Islands, in the Seychelle Islands.
Remarks. The individual specimens are noted as follows:
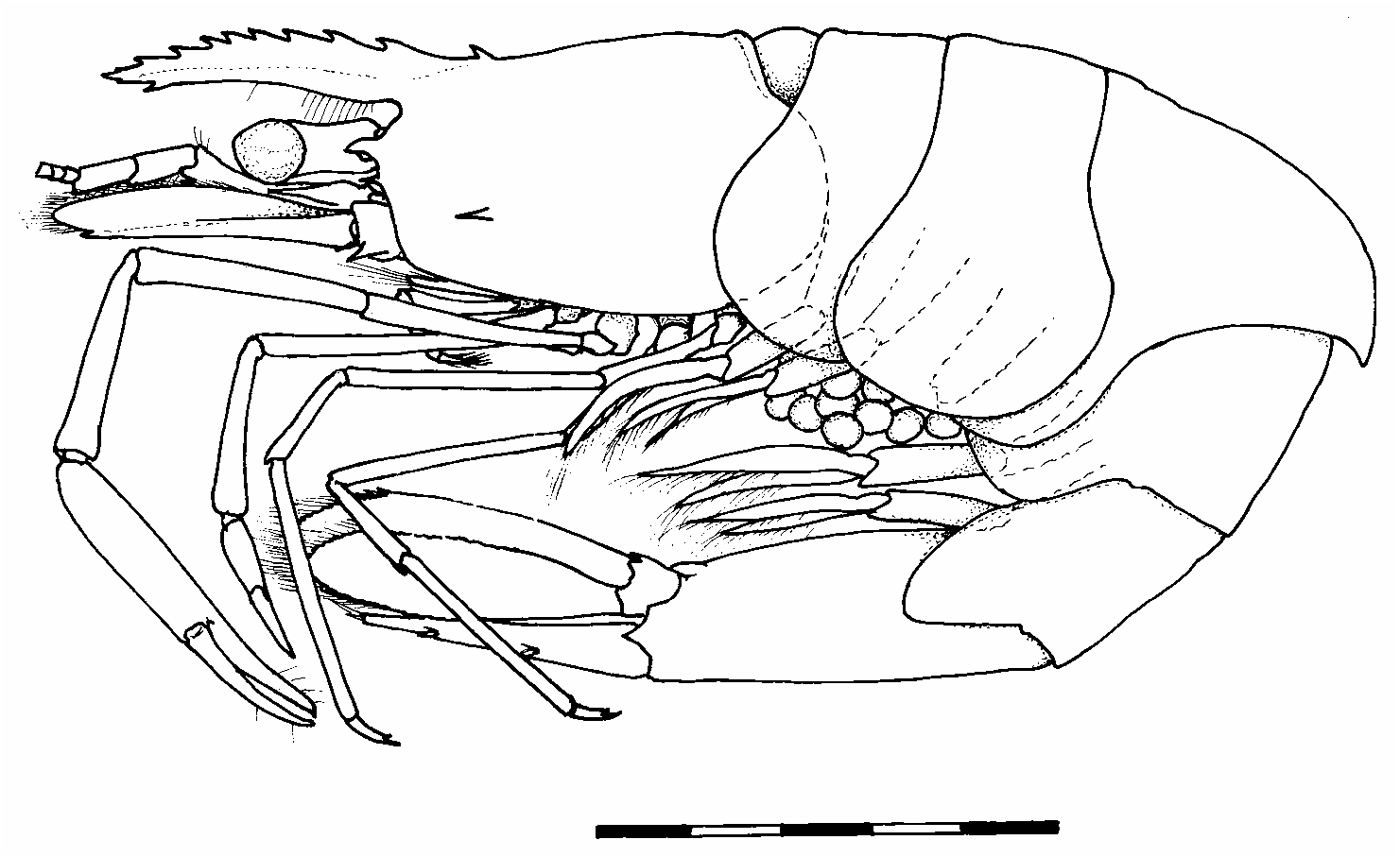
(1A) Ovig. ♀, holotype ( Figs 1–2 View FIGURE 1 View FIGURE 2 ), CL 3.9 mm, R. 1+9/1(+1), is complete except for right second pereiopod and left fifth pereiopod. Second pereiopod with palm 1.03 times carpal length. Cornea appears poorly pigmented, whitish, in preserved specimen .
(1B) Ovig. ♀, paratype ( Fig. 3 View FIGURE 3 ), CL 3.7 mm, R.1+8/2+(1), with carapace detached, with antennae and eyes, thoracic sternites and pereiopods attached, both second pereiopods attached. Second pereiopod with palm 1.03 and 1.10 times carpal length .
(1C) Ovig. ♀, paratype, CL 3.5 mm, R.1+8/2, with carapace almost completely detached from thorax, abdomen detached, most mouthparts in situ, with right first pereiopod attached, without second pereiopods, with right third and fifth pereiopods attached. Generally as holotype, but distal ventral rostral tooth better developed .
(1D) Ovig. ♀, paratype, CL 2.9 mm, R. 1+9/2+(1), with carapace detached, with eyes and left antennae only; without thoracic sternites or attached pereiopods; abdomen with sixth segment and caudal fan detached, telson lacking. Left antenna detached, acute proximo-dorsal process of basicerite apex angulate anteriorly. One second pereiopod attributed to this specimen. Second pereiopod with palm 0.90 of carpal length
(2) The single specimen from Malé ( Fig. 4 View FIGURE 4 ) is also incomplete, without eyes and right scaphocerite, with both first pereiopods but without second pereiopods (one missing, one small regenerating); with the right fourth pereiopod only; and with the sixth abdominal segment and caudal fan detached from the fifth. The proximal segment of the antennular peduncle with proximo-dorsal process.
Only four second pereiopods are preserved and in all of these, the carpus is about as long as the palm of the chela. In the aesopius- group, only P. longicarpus Bruce and Svoboda, 1983 , and P. speciosus have the second pereiopod carpus distinctly longer than the palm of the chela, the former species is immediately distinguished from P. amirantei by the absence of any post-rostral carapace spines or teeth.
One mini-tube of appendages contains a first and a second pleopod, but it is not clear from which specimen they were detached. The endopod of the first pleopod bears a well developed medial accessory lobe and the distolateral margin has several short plumose setae. The endopod of the second pleopod bears a subcylindrical appendix masculina with four similar simple terminal spines, about subequal to the corpus length. The appendix interna is subequal to the appendix masculina corpus length.
Another tube contains a telson, presumably from specimen 1D.
Periclimenes amirantei also shows a close resemblance to the specimen from Banda, Siboga station 240, reported by Holthuis ( Periclimenes (Periclimenes) aesopius, 1952: 34 –37 partim, fig. 6). This specimen, Ε, CL 4.6mm, has a rostral dentition of 1+8/2, with minute distal and ventral teeth and concave ventral margin. The hepatic spine is distinctly anterior to the level of the epigastric spine. The third abdominal tergite is feebly and bluntly posteriorly produced, non-carinate. The second pereiopod carpus is shorter than the palm length, about 0.85. The ambulatory propods and dactyls appear similar.
Urocaris longicaudata has also been reported from 9.0 metres at Aripu Paar, Sri Lanka, by Pearson (1905). It has not proved possible to locate the single ovigerous female specimen. Holthuis (1952) referred the specimen to Periclimenes aesopius (Bate) . The specimen has an arched rostrum with a dentition of 1+8/2 ( Pearson, 1905, Pl. XLIV, figs 5A, 5b) which closely resembles a typical aesopius group rostrum. Pearson unfortunately does not describe or illustrate the morphology of the pereiopods and it is therefore not possible to identify the specimen to species level. It was not included in the catalogue of material from that expedition held by Liverpool University in the 1950’s and is not in the collections of the World Museum Liverpool (Dr Ian Wallace, pers. com., 7-3-07). It is probably no longer extant.



| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.