
Ameira kimchi, Karanovic, Tomislav & Cho, Joo-Lae, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.281703 |
|
DOI |
https://doi.org/10.5281/zenodo.6171001 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE87BD-3E16-6F52-FF30-61CEC10997F3 |
|
treatment provided by |
Plazi |
|
scientific name |
Ameira kimchi |
| status |
sp. nov. |
Ameira kimchi sp. nov.
( Figs. 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Type locality. South Korea, South Sea, Memuld Island, littoral, depth 33 m, 34.654683°N 128.593633°E.
Specimens examined. Types only: holotype ovigerous female dissected on one slide (collection number NIBRIV0000232635), one paratype ovigerous female in toto on slide (collection number NIBRIV0000232636), one paratype ovigerous female dissected on one slide (collection number NIBRIV0000232637), and allotype male dissected on one slide (collection number NIBRIV0000232638); all collected from type locality, 18 December 2010, temperature 15.1 °C, salinity 33.84 psu, pH 7.54, leg. W. Lee.
Etymology. The species in named after kimchi , traditional Korean food, which is a range of fermented vegetables (most usually cabbages) consumed today by most Koreans on a daily basis. Senior author’s fondness for kimchi , even before living in Korea, contribute to a view of this dish as an embodiment of the traditional Korea. The name should be treated as a Latin noun in apposition.
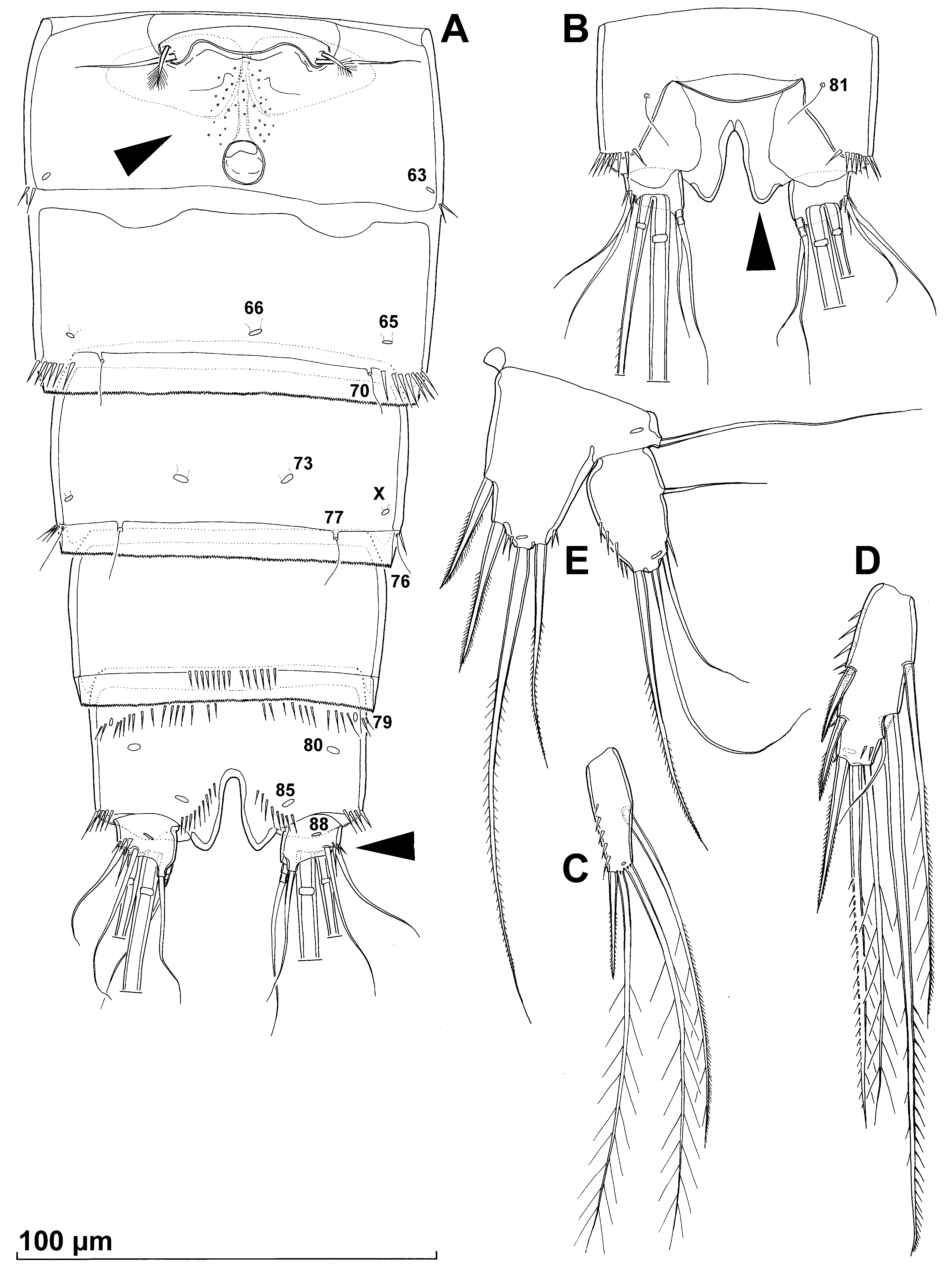
Description. Female (based on holotype and two paratypes). Total body length, measured as in previous species, from 442 to 452 μm (445 μm in holotype). Preserved specimen yellowish. Nauplius eye not visible. Body segmentation as in previous species. Most somite ornamentation also similar to previous species, and presumably homologous pore and sensilla also numbered with same Arabic numerals (see Figs. 5 View FIGURE 5 A, B, 6A, B) to allow easier comparison. Habitus ( Fig. 5 View FIGURE 5 A, B) more or less cylindrical, and generally tappering towards posterior end, with clear constriction on podoplean boundary between prosome and urosome in dorsal view; prosome/urosome ratio only 0.9 but greatest width in dorsal view also at posterior end of cephalothorax as in previous species. Body length/width ratio about 3.8 in dorsal view; cephalothorax 1.2 times as wide as genital double-somite. Free pedigerous somites without lateral or dorsal expansions, pleural plates only partly covering coxae of swimming legs in lateral view. Integument relatively strongly chitinized and smooth, without cuticular windows or pits except small area between genital aperture and copulatory pore (arrowed in Fig. 6 View FIGURE 6 A). Surface ornamentation of somites consisting of 83 pairs and seven unpaired pores and sensilla (those homologous with previous species indicated with Arabic numerals in Figs. 5 View FIGURE 5 A, B, 6A, B; those not present in previous species numbered with Roman numerals consecutively from anterior to posterior end of body, and from dorsal to ventral side), and several rows of spinules on urosomites only. Unpaired dorsal pore no. 17 homologous to sensillum no. 17 in previous species. Rostrum similar to previous species in shape and size, and also ornamented with two dorsal sensilla (pair no. 1).
Cephalothorax ( Fig. 5 View FIGURE 5 A, B) gradually tapering from 3/4 of its length towards anterior end in dorsal view, about 1.2 times as long as wide; represents 30% of total body length. Surface of cephalic shield ornamented with three pairs of lateral pores (nos. I, II, 14), two unpaired dorsal pores (nos. IV, 17), one unpaired dorsal sensillum (no. V), and 21 pairs of long sensilla (nos. 2–10, 12, 13, 15, 16, 18–21, 23–26,); sensilla 19–26 & VI belong to first pedigerous somite incorporated into cephalothorax; pairs of sensilla nos. 11 and 22 observed in previous species missing. Second pedigerous somite (first free) without pair of pores antero-laterally (no. 28 in previous species), ornamented with nine pairs of long sensilla (nos. 27, 29–35, VII); antero-dorsal pair of sensilla (no. 27) serially homologous to pair no. 19 on first pedigerous somite. Third pedigerous somite ornamented similarly to second one (also without antero-lateral pair of pores, no. 37 in previous species), with nine pairs of long sensilla (nos. 36, 38–45); antero-dorsal pair of sensilla (no. 36) serially homologous to pairs nos. 19 and 27 on previous two pedigerous somites. Fourth pedigerous somite also missing antero-dorsal pair of pores (no. 46), as in previous two somites, ornamented with seven pairs of long posterior sensilla (nos. 47–53). Hyaline fringes of all prosomites braod and smooth, except on fourth pedigerous somite where fringe very narrow dorsally. Fifth pedigerous somite ornamented with four pairs of posterior sensilla (nos. 54–57), as well as one pair of lateral pores (no. 58); hyaline fringe smooth and narrow.
Genital double-somite ( Figs. 5 View FIGURE 5 A, B, 6A) 0.9 times as long as wide (ventral view); internal suture (remnant of segmental fusion) very strongly sclerotised, wide, visible all around somite, furnished with two parallel short rows of spinules (two dorsal and two lateral) which much longer than in previous species, four pairs of sensilla (nos. 59–62), and two lateral pairs of cuticular pores (nos. 63, VIII); posterior part of genital double-somite ornamented with two pairs of large lateral pores (nos. 64, 65), one unpaird ventral pore (no. 66), one unpaired dorsal pore (no. IX), posterior row of spinules on eash side laterally (also longer than in previous species), and four pairs of posterior sensilla (nos. 67–70); hyaline fringe finely serrated. Genital complex ( Fig. 6 View FIGURE 6 A) with single large copulatory pore, weakly sclerotized and narrow stright copulatory duct (tapering towards anterior part), and two large triangular seminal receptacles. Single median genital aperture covered by fused reduced sixth legs, represents 41% of somite’s width. Integument between genital aperture and copulatory pore covered with shallow cuticular pits (arrowed in Fig. 6 View FIGURE 6 A). Third urosomite ornamented with posterior row of spinules dorsolaterally (narrowly interrupted dorsally), with unpaired dorsal cuticular pore (no. 71), three pairs of cuticular pores (nos. 72, 73, X), unpaired dorsal posterior sensillum (no. 74), and three pairs of posterior sensilla (nos. 75–77); hyaline fringe finely serrated. Fourth urosomite ornamented with single pair of lateral cuticular pores (no. 78), and short posterior row of slender spinules ventrally; hyaline fringe finely serrated. Anal somite ( Figs. 5 View FIGURE 5 B, 6A, B) clefted medially at posterior third, with median posterior margin on both sides of cleft strongly sclerotised and produced posteriorly (arrowed in Fig. 6 View FIGURE 6 B), with pair of large dorsal sensilla (no. 81), seven pairs of cuticular pores (nos. 79, 80, 82, 84, 85, XI), ventro-lateral anterior row of slender spinules, dorso-lateral posterior row of spinules at base of each caudal ramus, and row of slender spinules ventrally posterior to most median cuticular pore (no. 85); pairs of pores nos. 83, 84, 86, 87 missing (present in revious species); anal operculum convex, wide and short, reaching only proximal half of anal somite, represents 47% of somite's width, smooth; anal sinus also completely smooth, widely open.
Caudal rami ( Figs. 5 View FIGURE 5 A, 6A, B) extremely small compared to anal somite (arrowed in Fig. 6 View FIGURE 6 A), only about 0.25 times as long as anal somite, about 0.8 times as long as wide (ventral view), parallel and nearly cylyndrical, with space between them twice that of one ramus width, and with dorsal sclerotised side very short; with seven elements (three lateral, one dorsal and three apical); ornamentation consists of two spinules at base of both large lateral setae, and single ventral pair of pores (no. 88); dorsal piar of pores (no. 87) missing. Dorsal seta long and slender, smooth, inserted on postero-median corner, about 3.7 times as long as caudal ramus, triarticulate at base. Lateral setae all smooth and slender, all inserted very close to each other and close to posterior margin; ventralmost seta (ancestral distal) longest, more than five times as long as minute middle seta (ancestral smaller proximal), 1.3 times as long as dorsalmost seta (ancestral larger proximal), about as long as ventral seta, and about 3.3 times as long as caudal ramus. Inner apical seta smooth, about as long as ventralmost lateral seta. Middle apical seta strongest, with breaking plane, finely bipinnate at middle third of its length, almost as long as all free (postcephalothoracic) somites combined. Outer apical seta also with breaking plane and strong, finely unipinnate along outer margin, about 0.4 times as long as middle apical seta.
Antennula ( Fig. 5 View FIGURE 5 B) eight-segmented, joined to cephalotholax with small triangular pseudosegment laterally, approximately 0.8 times as long as cephalothorax, unornamented, armed as in previous species.
Antenna ( Fig. 5 View FIGURE 5 C) armed and ornamented as in previous species, but exopod missing enlarged spinule, and its other spinules much more slender.
Labrum ( Fig. 5 View FIGURE 5 B) large compared with cephalothorax, trapezoidal, rigidly sclerotized, with relatively short and somewhat concave cutting edge, ornamented subapically with two rows of eight strong spinules and apically with minute spinules. Also not mounted in satisfactory position to allow independent drawing
Paragnaths very similar to previous species.
Mandibula ( Fig. 5 View FIGURE 5 D) with coxa very similar to previous species. Palp with additional large spinule at base of basal seta, and also with basis more inflated distally; lateral endopodal seta longer and inserted more proximally than in previous species.
Maxillula and maxilla without any difference from those in previous species.
Maxilliped ( Fig. 5 View FIGURE 5 E) segmentation and armature as in previous specis; syncoxa with fewer and smaller spinules; basis with shorter spinules on inner margin, and one additional distal row of long and slender spinules on outer margin; endopodal claw slightly proportionately longer than in previous species.
All swimming legs ( Figs. 5 View FIGURE 5 F, 6C, D) of similar size and with same segmentation, armature, and ornamentation as in previous species.
First swimming leg ( Fig. 5 View FIGURE 5 F) with smooth intercoxal sclerite, its distal margin slightly concave. Praecoxa small and triangular, ornamented with row of small spinules on anterior surface along distal margin. Coxa 1.6 times as wide as long, with cuticular pore on anterior surface close to inner margin, and with two parallel rows of spinules along outer margin, which shorter than in previous species. Basis with longer and stronger spine on outer corner than in previous species (also finely bipinnate), and one finely bipinnate strong spine on inner distal corner, which also longer than in previous species; ornamented with spinules at base of each spine, as well as with distal row of spinules between exopod and endopod, and cuticular pore on anterior surface close to outer spine; inner margin smooth. Exopod as in previous species, except for smooth inner margin of third segment. Endopod geniculate and 1.6 times as long as exopod; first endopodal segment as long as exopod and 4.2 times as long as wide, ornamented with slender spinules along inner margin and two spinules along distal margin, with single strong and finely bipinnate inner seta, which about 0.7 times as long as segment; second segment smallest, about 1.5 times as long as wide and only 0.25 times as long as first segment, ornamented with three spinules on outer distal corner, and with single slender and sparsely bipinnate seta on inner distal corner; third segment slender, about four times as long as wide and nearly twice as long as second segment, armed apically with three elements; innermost apical element probably spine, strong and 1.6 times as long as third segment, unipinnate along outer margin; middle element more slender and geniculate, twice as long as outer element, and finely unipinnate along outer margin distally; innermost element slender bipinnate seta, 0.4 times as long as outer element.
Second swimming leg ( Fig. 6 View FIGURE 6 C), third swimming leg, and fourth swimming leg ( Fig. 6 View FIGURE 6 D) same as in previous species, except for inner seta on third endopodal segment of second leg proportionately longer (compare Figs. 3 View FIGURE 3 B and 6C), as well as third exopodal segment of fourth leg with longer distal inner seta and shorter inner apical seta (compare Figs. 3 View FIGURE 3 D and 6D).
Fifth leg ( Fig. 6 View FIGURE 6 E) segmentation, armature and ornamentation very similar to that in previous specis, except endopodal lobe more elongated (reaching 2/3 of exopod) and additionally ornamented with four spinules along distal margin; most spiniform elements not as strong as in previous species.
Sixth legs ( Fig. 6 View FIGURE 6 A) completely fused together, indistinct, forming simple operculum covering single gonopore, without any ornamentation, each with two setae; inner seta smooth, directed laterally, and about three times as long as outer seta, which bipinnate distally and directed postero-laterally.
Male (allotype). Body length 430 μm. Habitus shape, colour, ornamentation of almost all somites and caudal rami ( Fig. 7 View FIGURE 7 A, B), rostrum, antenna, labrum, paragnaths, mandibula, maxillula, maxilla, maxilliped, second swimming leg, third swimming leg, and fourth swimming leg similar to female. Prosome/urosome ratio 1.1, greatest width at posterior end of cephalothorax, body length/width ratio about 4.1; cephalothorax 1.3 times as wide as genital somite. Genital somite and first abdominal (third urosomite) not fused.
Genital somite ( Fig. 7 View FIGURE 7 A, B) 1.7 times as wide as long, with large and longitudinally positioned spermatophore visible inside on left side, which longer than somite and 3.6 times as long as wide, its distal part reaching proximal half of third urosomite.
Thrid urosomite ( Fig. 7 View FIGURE 7 A, B) with posterior row of large spinules continuous ventrally (between ventral pair of sensilla; pair no. 70), but without pair of lateral pores no. 65 (arrowed in Fig. 7 View FIGURE 7 A, B).
Fourth urosomite ( Fig. 7 View FIGURE 7 A, B) with posterior continous row of large spinules ventrally (between ventral pair of sensilla; pair no. 77).
Anal somite ( Fig. 7 View FIGURE 7 A, B) shape and ornamentation as in female, also with produced and strongly sclerotised inner median corners at base of cleft (arrowed in Fig. 7 View FIGURE 7 B).
Caudal rami ( Fig. 7 View FIGURE 7 A, B) shape, proportions, armature and ornamentation as in female.
Antennula ( Fig. 7 View FIGURE 7 C) long and slender, eight-segmented, strongly digeniculate, with geniculations between third and fourth and between sixth and seventh segments; ornamented with short row of proximal spinules on first segment. Seventh segment with inner margin strongly chitinized, with several smaller and three larger spine-like structures. Very long and slender aesthetasc on fourth segment fused basally to equally long seta (both about as long as entire antennula when completely closed), and much shorter and more slender aesthetasc on eighth segment apically also fused to two apical setae. First two segments similar to female, and last segment homologous to last two in female. Setal formula: 1.9.7.9.1.2.1.11. Most setae smooth and slender; seta on first segment bipinnate; one seta on fourth and one on sixth segment very short, spiniform and unipinnate; all these setae distally slender and smooth and most with small pore on tip. Only five lateral setae on eighth segment barticulated basally (inserted on small pseudojoint). No setae with breaking plane.
First swimming leg ( Fig. 7 View FIGURE 7 D) with smooth and distally somewhat inflated modified inner spine on basis, about as long as basis and directed distally; inner margin of basis smooth.
Fifth legs ( Fig. 7 View FIGURE 7 A, B, E) smaller than in female and with baseoenopods fused medially, with similar shape and ornamentation to that in male of previous species. Endopodal lobe broad, convex, not extending to middle of exopod, unornamented (except for medial cuticular pore), with inflated inner distal margin, with only two elements: inner unipinnate spiniform element slightly longer than baseoendopod and about 3.6 times as long as outer slender and smooth seta. Exopod without spinules, shorter than in female, 1.2 times as long as wide, but with additional minute inner seta (which longer than in previous species), without third seta from inner side (so number of elements same as in female, but setae not homologous); spiniform bipinnate seta much longer than in previous species.
Sixth legs ( Fig. 7 View FIGURE 7 A, B) partly fused medially to each other and right leg completely fused basally to somite; left leg articulated basally and movable, forming effective flap; each leg with three smooth setae and with single pore on anterior surface; length ratio of setae, from inner side, 1: 3.3: 1.8.
Variability. Only one male and three females were collected and examined under compound microscope. No variable features or asymmetries were observed, and most differences between male and female specimens are all part of a normal sexual dimorphism in this family. It is unclear at this stage if the absence of the cuticular pore no. 65 in male (arrowed in Fig. 7 View FIGURE 7 B) is also part of sexual dimorphism or is it intraspecific variability, because only one male was studied. The same dilemma remains in regard to the posterior rows of spinules on the ventral surface of the third and fourth urosomites in male (compare Figs. 6 View FIGURE 6 A and 7B), but this is more probably sexual dimorphism, as it has been observed previously in some species of this family (see, for example, Karanovic 2004, 2006)
Remarks. Although the spine formula of the swimming legs and the fifth leg, as well as the relative size of the first endopodal segment of the first leg, are no different from those of the previous species, or from the above mentioned cosmopolitan A. parvula , A. kimchi sp. nov. differs from all described species of the genus Ameira by its extremely small caudal rami (arrowed in Fig. 6 View FIGURE 6 A), and rigidly sclerotised and posteriorly produced inner median corners of the anal somite (arrowed in Fig. 6 View FIGURE 6 B). The latter character has not, in fact, been reported for any member of the family Ameiridae , and produced inner median corners in some other harpacticoids are usually a result of enlarged spinules, never of the somite’s margin itself. These structures in A. kimchi are so well sclerotised that they are observable even under a dissecting miscroscope.
A close examination of A. kimchi and A. zahaae showed that these two species also differ markedly in a number of microcharacters, previously not normally studied in detail in this group of crustaceans. Here we primarily refer to the pores and sensilla pattern, but some others are even more obvious, equally useful, and also very stable. For example, in A. kimchi the mandibular basis is much wider distally, with an additional large spinule; the first maxillipedal segment is much less ornamented; the genital somite has a field of cuticular pits between the genital aperture and copulatory pore (arrowed in Fig. 6 View FIGURE 6 A); the third urosomal somite in female bears no ventral spinules; the habitus is constricted in dorsal view between prosome and urosome, etc. As for the sensilla and pores pattern, when compared to A. zahaae , A. kimchi has 11 novel pores or sensilla (indicated with Roman numerals in Figs. 5 View FIGURE 5 A, B, 6C), nine pairs of sensilla or pores (nos. 11, 22, 28, 37, 46, 83, 84, 86, 87) are missing, and the unpaired ornamentation element no. 17 is expressed as a pore, not as a sensillum. We hope these and similar morphological comparisons will be used more widely in the future, as they hold a potential to resolve some dilemmas about dimorphism in this genus, such as that postulated for A. longispina Gee, 2009 with two distinct lengths of the inner basal spine on the male first leg (see Gee 2009). They may also show potential in distinguishing closely related species in species-complexes, as some molecular markers do ( Karanovic & Cooper 2011).
Small caudal rami are an unusual feature in the genus Ameira , and the only species with somewhat reduced caudal rami (although not as much as in A. kimchi ) is A. divagans Nicholls, 1939 , as redescribed by Scheibel (1974) from Germany. The two species, however, differ markedly in many characters. As far as we can judge based on morphology, A. kimchi has no close relatives among living congeners, and its place in the A. parvula speciescomplex is almost entirely based on plesiomorphic characters (see the Remarks section for A. zahaae above).
Genus Pseudameira Sars G.O., 1911



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |