
Deontostoma tridentum, Leduc, Daniel, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3722.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:C5B74DF2-72F1-4C77-A9D4-37AB7B06B4FD |
|
DOI |
https://doi.org/10.5281/zenodo.5691579 |
|
persistent identifier |
https://treatment.plazi.org/id/4A6743DA-EB31-46DE-B8E4-EF4F422DF552 |
|
taxon LSID |
lsid:zoobank.org:act:4A6743DA-EB31-46DE-B8E4-EF4F422DF552 |
|
treatment provided by |
Plazi |
|
scientific name |
Deontostoma tridentum |
| status |
sp. nov. |
Deontostoma tridentum n. sp. ( Table 1 View TABLE 1 , Fig. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Material examined. Holotype male (NIWA 88367), collected on 11 November 2011 (NIWA cruise TAN1116, station 68), southern flank Chatham Rise (44.221° S, 178.882° E), water depth: 1007 m, sediment total organic matter content (0–5 cm): 2.9%, sediment CaCO3 content (0–5 cm): 21.8%. Three paratype males and two paratype females (NIWA 88368), same data as holotype. One paratype female (NIWA 88369), collected on 6 November 2011 (NIWA cruise TAN1116, station 33), southern flank Chatham Rise (43.389° S, 174.175° E), water depth: 570 m, sediment total organic matter content (0–5 cm): 2.7%, sediment CaCO3 content (0–5 cm): 7.8%.
Etymology. The species name is derived from the Latin tres (= three) and dens (= tooth), and refers to the presence of three onchia in the buccal cavity of this species.
Description. Male. Body large, cylindrical, gradually tapering from mid–body to anterior end. Cuticle thick, thinnest in anterior pharyngeal region (~14 µm) and gradually thickening posteriorly (up to ~19–21 µm thick), comprising several distinct layers as seen under light microscope ( Figure 2 View FIGURE 2 A); light transverse striations anterior to amphid and oblique striations crossing at angle of ca 120 ° in anterior pharyngeal region ( Figure 2 View FIGURE 2 B and 3D), smooth elsewhere. Head blunt, rounded, set-off by slight constriction at level of amphideal aperture. Six small inner labial papillae with conspicuous ducts at base; six outer labial setae and four cephalic setae in one circle, equal in length, 10–12% cbd long, with nerve process often visible. Eight rows of somatic setae (two subventral, two subdorsal, two lateral, one ventral and one dorsal) in anterior pharyngeal region, sometimes in clusters of two or three, with conspicuous ducts; somatic setae short and sparse elsewhere. Cephalic capsule without prominent anterior lobes or tropis; six posterior lobes present. Interlobar incisions wide, simple, without posteriorly-directed arms; posterior lobe margin smooth, with shallow incisions. Intralobar lacunae mostly absent. Amphids pocketshaped with relatively small aperture, 16–17 µm wide by 3–5 µm high.
Species Deontostoma tridentum n. sp.
Males Females Mouth opening wide, triangular (en face view), enclosed by one dorsal and two subventral microlabia ( Figure 3 View FIGURE 3 A and B). Subventral microlabia completely divided by mandibular ridge; dorsal microlabium only partially divided by mandibular ridge ( Figure 4 View FIGURE 4 C). Odontia reduced to minute ridges and not visible with light microscopy ( Figure 3 View FIGURE 3 B), appear completely absent in some specimens ( Figure 4 View FIGURE 4 A). Buccal cavity with large dorsal onchium and two smaller subventral onchia. Pharynx widening gradually posteriorly, ocelli not observed. Nerve ring at 22% of pharynx length from anterior. Pharyngeal glands and secretory-excretory system not observed.
Male reproductive system diorchic with opposed testes. Two rows of subventral pre-cloacal supplements consisting of 14–17 setae beginning slightly posteriorly to cloaca followed by 6–7 pairs of papilloid supplements bearing setae ( Figure 2 View FIGURE 2 E and 3C). Transition between setae and papillae bearing setae is gradual and distinction between the two not always clear. Distance between papilloid supplements increases anteriorly from ca 60 to 120 µm. One ventral supplement situated 200–215 µm anterior to cloaca, consisting of a central oval projection flanked by two lateral ridges with rounded outline; central projection with minute opening ( Figure 3 View FIGURE 3 E). Spicules paired, slightly curved, 1.2–1.4 abd long, with central lamina extending along almost entire length of spicules.
Gubernaculum with broad proximal ends without ornamentation and curving laterally; triangular crurae extending from middle portion. Proximal portion of gubernaculum is rounded, appears to consist of glandular tissue surrounded at base by cuticularised portion of gubernaculum. Copulatory musculature well-developed, extends ca 2000 µm anterior to cloaca.
Tail short, cylindrical, with blunt, rounded tip. Three caudal glands extending anterior to cloaca; spinneret present ( Figure 2 View FIGURE 2 C). Eight rows of short somatic setae present (two subventral, two subdorsal, two ventrosublateral, and two dorsosublateral).
Females. Similar to males, but with smaller amphids and slightly longer tails. Reproductive system amphidelphic with two opposed and reflexed ovaries situated to the left of intestine. Vulva situated at ~2/3 of body length, surrounded by small intracuticular granules ( Figure 2 View FIGURE 2 D). Up to seven mature eggs present, 250–290 µm wide by 735–810 µm long.
Diagnosis and relationships. Deontostoma tridentum n. sp. is characterised by large buccal cavity with greatly reduced or inconspicuous odontia, one large dorsal onchium and two smaller subventral onchia, cephalic capsule without interlobar incisions with posteriorly–directed arms, ventral pre-cloacal supplement situated 200–215 µm anterior to cloaca, and gubernaculum with triangular crurae and curved proximal portion.
Deontostoma tridentum n. sp. can be differentiated from all species of the genus based on the presence of two subventral onchia in the buccal cavity (absent in all other species). It is most similar to D. coptochilus in the presence of a large buccal cavity with a prominent dorsal onchium, structure of the cephalic capsule, and structure of the microlabia (i.e., subventral microlabia completely divided by mandibular ridge and dorsal microlabium not completely divided by mandibular ridge). Deontostoma tridentum n. sp. differs from the latter in the presence of crurae (absent in D. coptochilus ), absence of ocelli (present in D. coptochilus ), and in the position and structure of the ventral pre-cloacal supplement (> 200 µm anterior to cloaca, central longitudinal ridge absent in D. tridentum n. sp. vs <110 µm anterior to cloaca, central longitudinal ridge present in D. coptochilus ). Deontostoma tridentum n. sp. is also similar to D. parantarcticum in the presence of more than one onchia in the buccal cavity and absence of ocelli, but can be differentiated from the latter by smaller values of a (36–52 vs 60–73), longer cephalic setae (7–8 vs 3–4 µm), absence of conspicuous odontia (vs 2 large odontia on dorsal microlabium present in D. parantarcticum ), and presence of 6–7 subventral pre-cloacal papillae (vs 3 in D. parantarcticum ).
Remarks The Brenke sledge was primarily designed to sample organisms present in the benthic boundary layer, i.e., suprabenthic organisms (Brandt & Barthel 1995), but re-suspended benthic organisms are also often sampled (e.g., Lörz et al. 2011). Deontostoma tridentum n. sp. specimens were found in core samples obtained from the type localities (D. Leduc, unpublished data), which suggests that the presence of specimens in the sledge nets was due to re-suspension of sediments. Association of this species with larger organisms such as sponges could also have increased its chance of capture by the sledge nets, but an association of D. tridentum n. sp. with other organism(s) has yet to be verified.

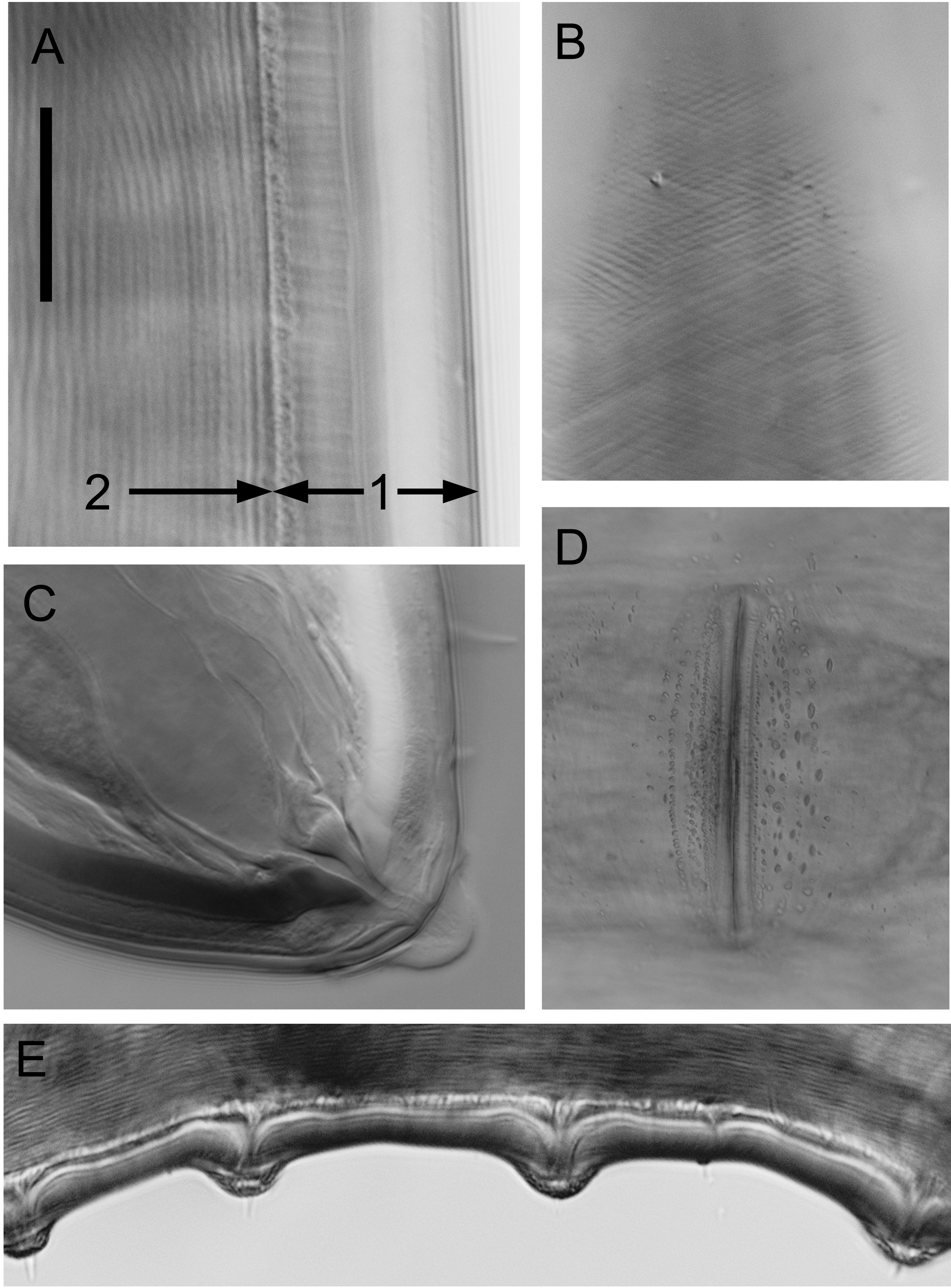
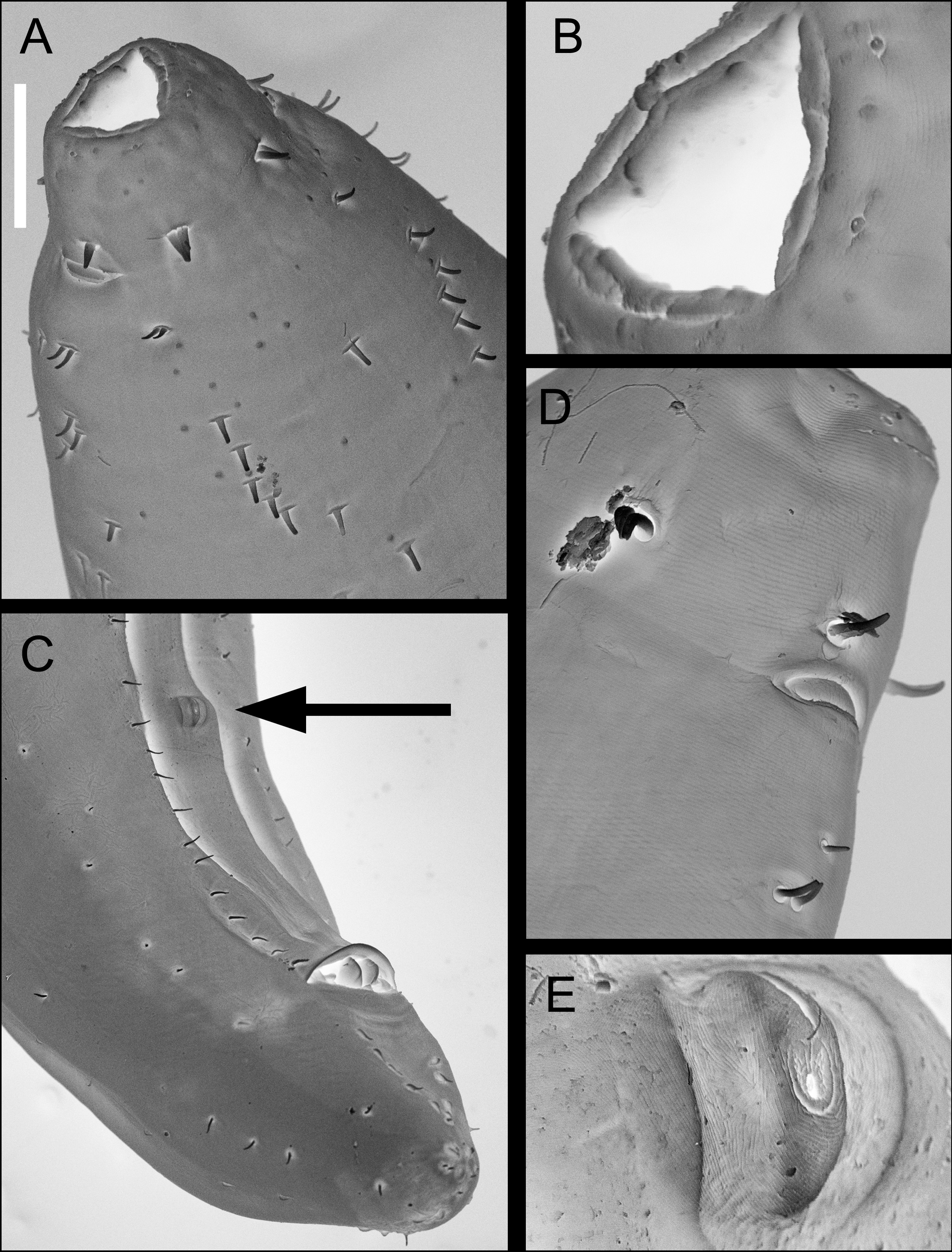

TABLE 1. Morphometrics (µm, unless specified otherwise) of Deontostoma tridentum n. sp. (a, body length / maximum body diameter; abd, anal body diameter; b, body length / pharynx length; c, body length / tail length; cbd, corresponding body diameter; V, vulva distance from anterior end of body; % V, V / total body length)
| Holotype | Paratypes | Paratypes | |
|---|---|---|---|
| n | – | 3 | 3 |
| L (mm) | 21.6 | 18.7–19.7 | 15.2–23.0 |
| a | 49 | 48–52 | 36–52 |
| b | 6 | 5–6 | 5–6 |
| c | 110 | 93–112 | 72–110 |
| Head diam. at level of amphids | 67 | 65–69 | 66–70 |
| Head diam. at level of ceph. setae | 58 | 59–64 | 63–65 |
| Mouth diam. | 13 | 10–13 | 11–14 |
| Length of cephalic setae | 7–8 | 7–8 | 7 |
| Amphid height | 13 | 12–13 | 9–14 |
| Amphid width | 17 | 16 | 10–12 |
| Amphid width/cbd (%) | 25 | 23 | 14–18 |
| Amphid from anterior end | 33 | 29–30 | 26–30 |
| Cephalic capsule height | 54 | 46–49 | 47–50 |
| Nerve ring from anterior end | 799 | 794–839 | 803–827 |
| Nerve ring cbd | 222 | 220–229 | 194–199 |
| Pharynx length | 3603 | 3378–3751 | 3079–3623 |
| Pharynx diam. at base | 133 | 136–165 | 127–146 |
| Pharynx cbd at base | 317 | 338–361 | 310–359 |
| Max. body diam. | 442 | 377–411 | 422–443 |
| Spicule length | 285 | 223–256 | – |
| Gubernacular apophysis length | 104 | 105–115 | – |
| Anal body diam. | 199 | 192–195 | 164–178 |
| Tail length | 196 | 167–203 | 197–212 |
| Tail length/abd | 1.0 | 0.9–1.1 | 1.2–1.3 |
| V (mm) | – | – | 9.3–14.8 |
| %V | – | – | 61–71 |
| Vulvar body diam. | – | – | 364–419 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |