
Grimpoteuthis imperator, Ziegler & Sagorny, 2021
|
publication ID |
https://doi.org/ 10.1186/s12915-021-01000-9 |
|
DOI |
https://doi.org/10.5281/zenodo.4725952 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB87C2-FFE1-FFD8-44AD-FDF8FB248683 |
|
treatment provided by |
Felipe |
|
scientific name |
Grimpoteuthis imperator |
| status |
sp. nov. |
Grimpoteuthis imperator View in CoL sp. nov.
( Figs. 1 View Fig and 2 View Fig ; Tables 2 View Table 2 and 3 View Table 3 ; Additional files 1, 2, and 3; [ 32])
Synonymy
Grimpoteuthis sp. ([ 16]: 83, Fig. 114A)
Grimpoteuthis sp. ([ 33]: 265; Fig. 10F, G)
Material examined
HOLOTYPE: mature male, ML 95 mm; R/V SONNE, SO-249 BERING, DR 82 ; 48° 27.53 ′ N, 168° 52.21 ′ E, 3913 – 4417 m depth; chain bag dredge; 5 July 2016; ZMB MOLL 240160 . GoogleMaps
Diagnosis
Medium-sized species with moderately long, lateral fins. Cirri short and suckers moderate. Gills compact with eight broad lamellae. Radula present, teeth homodont. Paired anterior and unpaired posterior salivary glands present. Shell U-shaped, smooth with lateral wings parallel, broadly tapering towards distal ends.
Description
Medium-sized species, body semi-gelatinous, bell-shaped ( Fig. 1a – e View Fig ). Mantle about one third of total length (MLI 32.8), saccular, gelatinous, nearly twice as long as wide, broadly rounded posteriorly. Head wider than mantle (HWI 23.8), no neck region visible ( Fig. 1f View Fig ). Pallial aperture narrow, enveloping base of funnel ( Fig. 1e View Fig ). Funnel long (FuLI 30.4), protruding far beyond mantle margin, distal third of funnel free ( Fig. 1b View Fig ). Funnel broader at the base than at the opening ( Fig. 1g View Fig ). Funnel organ of inverted V shape. Olfactory organs rounded, situated within the pallial aperture on either side of the funnel base ( Fig. 1g View Fig ). Fins moderately long (FSI 68.6, FLI 100.0), about half as wide as long ( Fig. 1b View Fig ). Fins situated laterally, between the eyes and apex of the mantle, but closer to the eyes. Posterior fin margin straight, less gelatinous than the rest of the mantle. Tips of fins broadly rounded. Anterior fin margin slightly convex, tapering posteriorly towards the base. Fin cartilage and fin muscles inserting in the posterior portion of the fins ( Fig. 1d View Fig ). Broad muscular base, attached to the shell. Towards the tips of fins, the fin cartilage becomes narrower, ending in a small tip about ¾ of fin length.
Arms semi-gelatinous, subequal, moderately long (ALI 63.4). The fourth arm on ventral side shortest (ca. 54% of total length), first arm on dorsal side longest (ca. 63% of total length), arm formula varying between left and right sides. The arms deeply set into the soft and fleshy primary web, no secondary web present ( Fig. 1c View Fig ). Web formula differing between right and left sides with sector Aor Bdeepest and sector Dor E shallowest, respectively. Web nodules large and rounded, located on the ventral side of the arms between suckers 33 and 34, uniting web to the ventral side of the arms ( Fig. 1c View Fig ). Beyond the nodule, the web extends only a few suckers further, becoming very narrow. Nodules absent on the dorsal surface, web extending nearly to the tip of each arm. No nodules present in sector A, so the web extends nearly to the tip of both adjacent arms. In sector E, two nodules present, so the web extends only a short distance beyond the nodules, thus rendering this sector one of the shallowest sectors and sector A one of the deepest. Suckers in a single row extending from the mouth opening to the tips of the arms ( Fig. 1c View Fig ). Arms with 68 – 74 suckers set into the oral surface, their apertures projecting freely. First five suckers very small, followed by suckers gradually increasing in size (MSDI 4.3). No enlarged suckers around the web nodule, no sign of hectocotylization ( Fig. 1c View Fig ). Suckers cylindrical, with short, narrow apertural rims ( Fig. 1h View Fig ). Simple sucker aperture, the base of suckers embedded into primary web. Cirri short, 1.0× maximum sucker diameter (MCLI 4.3), located on the oral surface of the arms between base of suckers ( Fig. 1h View Fig ). First cirrus between suckers 4 and 5. Cirri extending to the tips of the arms with no change in length along the arms.
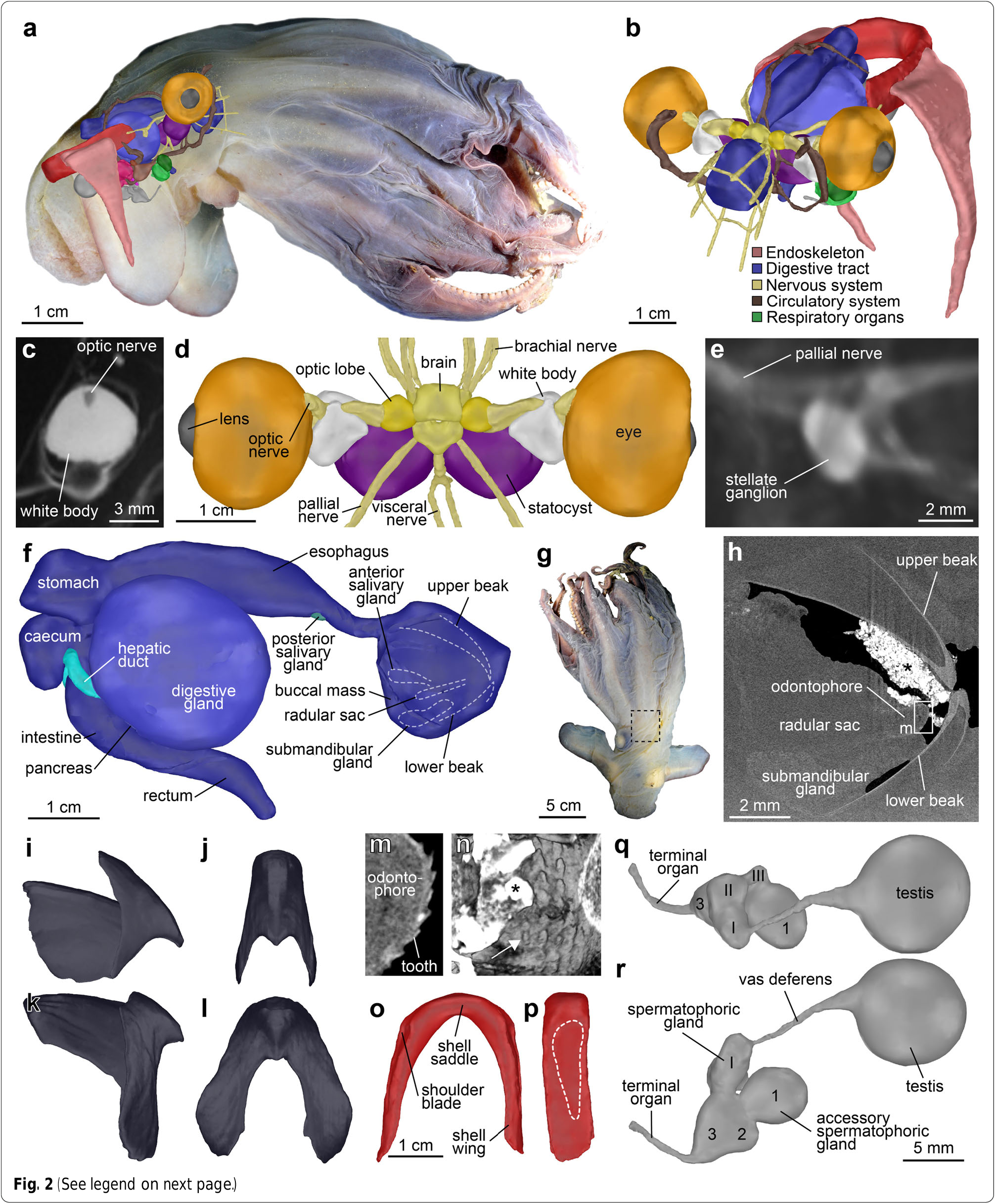
Gills compact (GDI 13.1), almost spherical ( Fig. 1i View Fig ), of ‘ half-orange ’ type, with eight broad lamellae each ( Fig. 1j View Fig ). Gills partly surrounding ovoid-shaped branchial glands. Teardrop-shaped branchial hearts located dorsoposterior to the gills, in close proximity to the renal appendages. Systemic heart with single large curved ventricle, indistinguishable auricles ( Fig. 1k View Fig ). Small swelling on the dorsal aorta directly dorsal to the ventricle. Dorsal aorta forming single large curve across the dorsal side of the oesophagus. Two large pallial arteries leaving the dorsal aorta as a single vessel each near the posterior end of the oesophagus. Eyes large (EDI 24.6), not protruding, with prominent lens ( Fig. 2a, b View Fig ). Optic lobes almost spherical in shape, with a single bundle of optic nerves passing through the white body ( Fig. 2c View Fig ) before tapering into several smaller nerves closer to the eye. White bodies more than twice the size of the optic lobes, somewhat spherical in shape with two processes directed towards the brain. Statocysts almost as large as the eyes ( Fig. 2d View Fig ). Stellate ganglion ovoid in shape, epistellar body morphologically not distinguishable ( Fig. 2e View Fig ).
The digestive tract in lateral view C-shaped with relatively large buccal mass ( Fig. 2f – h View Fig ). The upper beak without distinct folds, ridges, or thickenings ( Fig. 2i, j View Fig ). Lower beak with rounded hood and broad wings, lateral walls without any folds or ridges ( Fig. 2k, l View Fig ). Paired anterior salivary glands centrally located within the buccal mass, to the left and right side above the radular sac ( Fig. 2f View Fig ). Large median submandibular gland below radular sac ( Fig. 2f, h View Fig ). Radula poorly developed, with homodont dentition arranged in five rows ( Fig. 2m, n View Fig ). Rhachidian tooth slender, smaller than laterals. First lateral tooth broad triangular, second lateral tooth slender triangular, no trace of marginal teeth or plates. Oesophagus long, widening in the mid-part to form a simple crop ( Fig. 2f View Fig ). Unpaired posterior salivary gland small, located on the ventral side of the oesophagus. Stomach nearly rectangular, tapering towards the ventral side. The caecum slightly smaller than the stomach with two long and slender hepatic ducts leading to the large, unilobular, almost spherical digestive gland ( Fig. 2f View Fig ). Small pancreas present on the posterior surface of the digestive gland. Intestine shorter than the oesophagus, no enlargement. Rectum located close to the funnel. Anal flaps and ink sac absent.
Shell located in the dorsal part of the mantle cavity, robust, U-shaped ( Fig. 2a, b View Fig ). Shell wings not expanding, parallel to each other, broadly tapering towards the distal ends ( Fig. 2o View Fig ). Shoulder blades present, but weak. Outer surface of the shell saddle convex without median ridge or transverse groove. Fin cartilage insertions long, encompassing about ⅔ of the entire length of the shell wing ( Fig. 2p View Fig ).
Male reproductive system with large, almost spherical testis located centrally in the posterior part of the mantle, posterior to the stomach and caecum, ventral to the shell ( Fig. 2a View Fig ). Vas deferens elongated. Spermatophoric glands I – III moderately developed, convoluted ( Fig. 2q View Fig ). Accessory spermatophoric gland complex much larger than the spermatophoric gland complex. Accessory spermatophoric glands 1 and 3 nearly the same size, much larger than accessory spermatophoric gland 2 ( Fig. 2r View Fig ). Several spermatophores located inside the central duct within the accessory spermatophoric gland complex, each measuring ca. 2 × 1 mm. Terminal organ (penis) long and slender, directed towards the rectum ( Fig. 2r View Fig ).
Skin surface smooth. Dorsal surface of the head and mantle white, slightly reddish towards the posterior apex ( Fig. 1a View Fig ). Ventral surface of the head and mantle white with a red pigmentation, funnel darker red in colour, in particular at the distal end ( Fig. 1b View Fig ). Posterior margin of the fins deep red, becoming lighter and almost white towards the anterior margin. Dorsal arms on the surface white, dorsal surface of the web reddish ( Fig. 1a View Fig ). Ventral arms and web dark red, suckers lighter red ( Fig. 1c View Fig ).
Measurements and counts
Most morphometric and meristic characters were obtained from the holotype directly following capture ( Table 2 View Table 2 ).
Type locality
On a southwest-facing slope above a large circular basin southeast of Tenji Seamount, east of Winnebago Seamount, and northeast of Minnetonka Seamount, Emperor Seamounts, North Pacific Ocean; 48° 27.53 ′ N, 168° 52.21 ′ E, 3913 – 4417 m depth (https://tinyurl.com/rmsp7ej).
Distribution
So far known only from the type locality in the northern part of the Emperor Seamounts, an undersea mountain chain in the northwestern part of the North Pacific [ 34].
Remarks
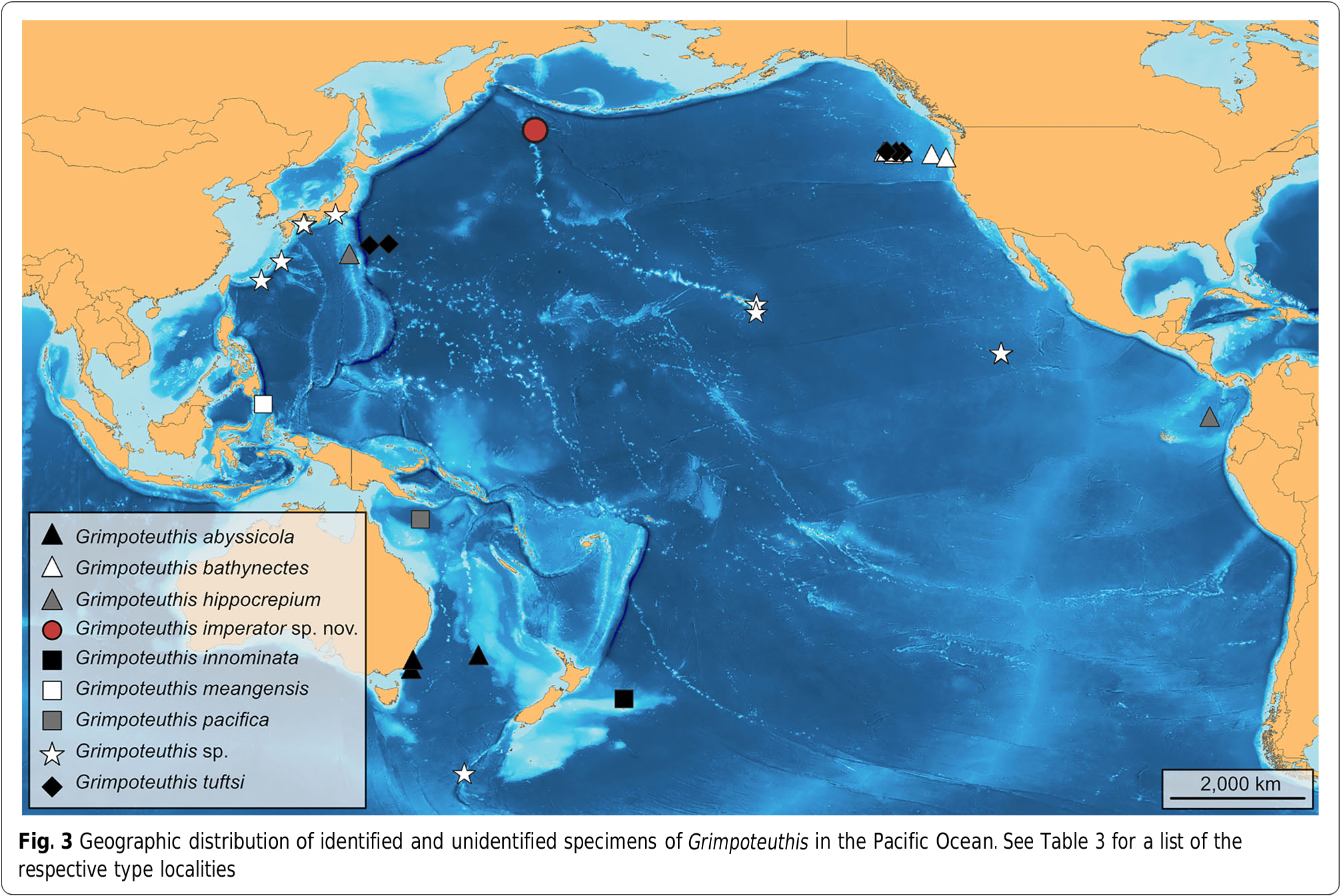
Based on the shell form, fin position, optic lobe shape, arm length, web form, and optic nerve arrangement, this specimen is readily identifiable as a cirrate of the genus Grimpoteuthis [ 14]. Differentiation from previously described species is based on the number of suckers (68 – 74), absence of enlarged suckers near the web nodules, absence of a thin web between the suckers, position of the web nodules (near suckers 33-34), cirrus length (1.0× maximum sucker diameter), location of proximal cirri (between suckers 4 and 5), presence of a radula, gill shape (‘ half-orange ’ type), gill lamellae count (8/8), and various shell characters ( Table 3 View Table 3 ). Apart from the new species described here, seven other Grimpoteuthis species as well as several unidentified specimens ascribed to this genus have been recorded from the Pacific Ocean ( Fig. 3 View Fig ). The type localities of the two geographically closest species, i.e. G. bathynectes and G. tuftsi , are also found in the North Pacific, but in areas more than 4000 km east of the type locality of G. imperator sp. nov. [ 30]. Further type localities of Pacific species pertaining to that genus are found in the tropical and southern parts of the ocean. Whilst only a single individual was analysed here, this is not uncommon in cirrate taxonomy due to the scarcity of suitable material [ 14] and under certain circumstances (e.g. ‘ unquestioned distinctiveness of the species ’ and ‘ the high probability that no additional material will be forthcoming soon ’) does not impede designation of a new species ([ 3]: 49).
Etymology
Latin, imperator, noun in apposition. Named after the Emperor Seamounts to which the type locality belongs. Proposed vernacular names are Emperor dumbo (English), Dumbo impérial (French), 天皇ダンボ (Japanese), and Kaiserdumbo (German).
DNA barcoding and phylogenetic analysis
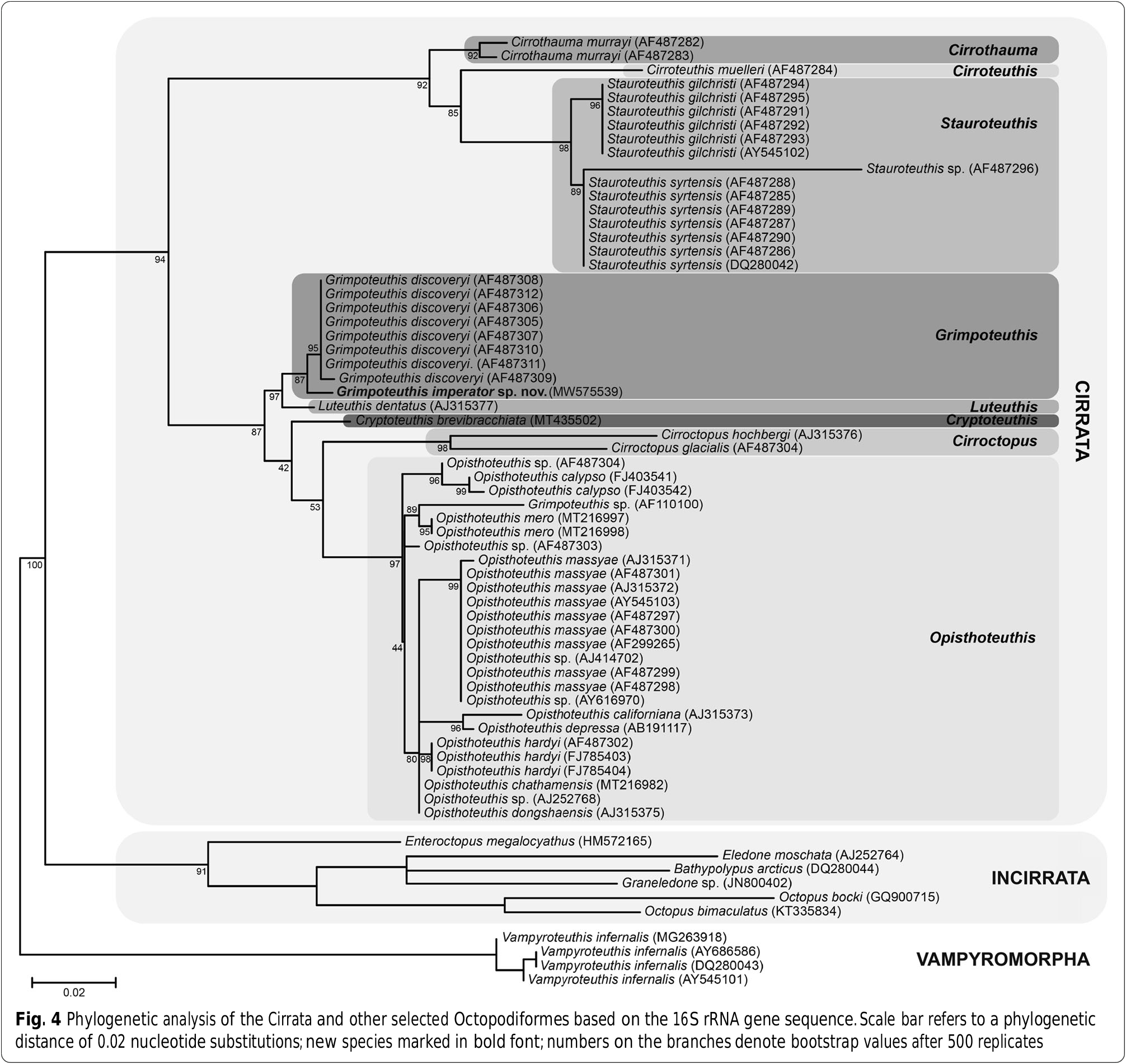
In addition to the non-invasively acquired morphological character states, minimally invasive DNA barcoding was used to obtain the first molecular sequence data for an unambiguously identified member of the genus Grimpoteuthis from the Pacific Ocean. Inferences based on this as well as 55 previously deposited 16S rRNA gene sequences result in the first phylogeny with full coverage of all eight extant cirrate genera ( Fig. 4 View Fig ).
The phylogenetic analysis provided very good resolution within and between cirrate genera as well as all outgroup taxa. The North Atlantic Grimpoteuthis specimens are all placed in a well-supported clade (95% bootstrap value). The new species G. imperator sp. nov. from the North Pacific is sister to this North Atlantic Grimpoteuthis clade (87%). Sister to all Grimpoteuthis species is Luteuthis dentatus , a branching that is statistically well-supported (97%). This larger clade in turn is sister to several species of the genera Cryptoteuthis , Cirroctopus , and Opisthoteuthis , a branching supported by a high bootstrap value as well (87%). A further, well supported (92%) clade composed of species from the cirrate genera Cirrothauma , Cirroteuthis , and Stauroteuthis is then sister to the clade composed of all previously mentioned cirrate taxa. Furthermore, all incirrate species included in the present analysis are sister to the well supported cirrate clade (94%). Finally, the vampire squid Vampyroteuthis is sister to all octopod species (100%).
Interspecific uncorrected p-distances based on the 16S rRNA gene fragment between G. imperator sp. nov. and Grimpoteuthis specimens from the North Atlantic vary between 2.2 and 2.5%. Interspecific distances between G. imperator sp. nov. and L. dentatus are 3.2% and between G. imperator sp. nov. and other cirrate species range from 8.9% ( Opisthoteuthis depressa ) to 16.6% ( Stauroteuthis syrtensis ). For reference, interspecific distances within other genera lie between 0.9% ( S. syrtensis and S. gilchristi ) and 4.3% ( O. hardyi and O. californiana ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Cirrata |
|
Family |
|
|
Genus |