
Vigtorniella ardabilia, Wiklund & Glover & Johannessen & Dahlgren, 2009
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2008.00469.x |
|
persistent identifier |
https://treatment.plazi.org/id/03EB87A8-BC29-1352-FC38-CEA73F193E95 |
|
treatment provided by |
Felipe |
|
scientific name |
Vigtorniella ardabilia |
| status |
sp. nov. |
VIGTORNIELLA ARDABILIA View in CoL SP. NOV.
( FIGS 2–4 View Figure 2 View Figure 3 View Figure 4 )
Type material: Northern North Atlantic , coastal Skagerrak, 58°53.1′N; 11°06.4′E, female with eggs, 6 mm long, 35 segments, preserved in formaldehyde from experimental tank with bone material sampled with ROV from an implanted Minke whale carcass at 125 m water depth, holotype (SMNHType-7376); same location, two specimens, one female and one male, preserved in formaldehyde, paratypes ( NHM2008.367 and 2008.368); same location, seven specimens preserved in formaldehyde, anterior parts of two specimens preserved in osmium for SEM, and several specimens preserved in ethanol for DNA extraction, all in first author’s collection. Northern North Atlantic , Svåsand in Hardangerfjord , 84 and 150 m depth, six specimens preserved in ethanol; northern North Atlantic, Mele in Hardangerfjord, 60°21.27′N; 6°20.89′E, 104 m depth, nine specimens preserved in formaldehyde GoogleMaps .
Description: Colour pale yellow in females with eggs ( Fig. 2 View Figure 2 ), males transparent, both with anterior five to six segments more reddish in live animals. Length up to 32 mm for 82 segments (male specimen). Body shape elongated, tapering slightly at anterior and posterior ends.
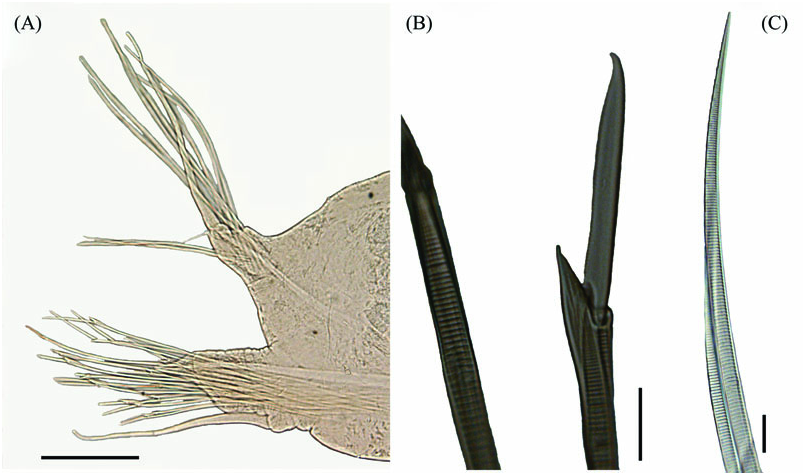
Prostomium ventrally displaced, not visible dorsally. Eye-pigment, four orange-red spots in a rectangular or trapezoidal arrangement, only visible in live animals. Median antenna and caruncle absent. Digitiform paired antennae inserted anteroventrally ( Fig. 3A View Figure 3 ). Palps spherical, partially fused, inserted ventrally directly anterior to mouth ( Fig. 3B View Figure 3 ). Retracted proboscis visible through epidermis, terminating posteriorly in segment 5–6. Jaws absent.
Segment 1 reduced, achaetous with only tentacular dorsal cirri, partially fused with segment 2. Segment 2 with dorsal and ventral tentacular cirri, notopodia, and neuropodia, with chaetae ( Fig. 3A, B View Figure 3 ). From segment 3, notopodia with dorsal cirri inserted distally, neuropodia with ventral cirri inserted basally ( Fig. 4A View Figure 4 ). Neuropodia in mid-body segments longer than notopodia. Notochaetae serrated spines, partly surrounding dorsal cirrus, with very fine serration not visible in light microscopy ( Figs 3E View Figure 3 , 4A, C View Figure 4 ). Neurochaetae finely serrated compound falcigers with laddered shafts (serration not visible in the light microscopy picture), the three to four upper neurochaetae with long slender blades, lower blades gradually shorter and wider ( Figs 3D View Figure 3 , 4A, B View Figure 4 ). Neurochaetae on segment 2 compound spinigers with thinner and longer serrations compared to more posterior segments ( Fig. 3C View Figure 3 ).
Pygidium with terminal anus and two pygidial cirri, unpaired appendage absent.
Distribution: Known from a whale-fall at 125 m depth in the Skagerrak, North Sea, and also found in sediment samples taken beneath fish farms in Hardangerfjord, Norway, at 80–150 m depth.
Etymology: Vigtorniella ardabilia sp. nov. is morphologically very similar to the ‘carpet-worm’ Vigtorniella flokati , and is named for the famous Ardabil Persian carpet, recently placed back on display at the Victoria and Albert Museum, London, UK.
Remarks: The species is morphologically almost identical to V. flokati , which was recorded in large aggregations (up to 8000 m-2) on a deep-sea whale-fall in the Santa Cruz Basin, California. One difference is that the antennae appear to be slightly longer on V. ardabilia compared to V. flokati ; on the former they are as long as the palps, on the latter they are less than half the length of the palps. A further difference is the presence of eyes in live specimens of V. ardabilia which are absent in V. flokati . As no live material of V. flokati was available for us to study, we cannot rule out the presence of eye spots in this species. Vigtorniella flokati have eggs in segments 6–9 only, whereas in V. ardabilia the eggs occur from segment 6 and backward in all segments to the end of the body. Vigtorniella ardabilia is different from V. zaikai in the presence of an achaetous tentacular segment (segment 1) which is not reported for V. zaikai , and in the absence of specialized whip-like chaetae and jaws. The neurochaetal spinigers that in V. ardabilia occur on segment 2 are not mentioned in the description of V. flokati ( Dahlgren et al., 2004) but must have been overlooked by the authors as they are present in their SEM micrograph (fig. 3A in Dahlgren et al., 2004). Vigtorniella flokati juveniles were found to have jaws, but jaws were not found in any of the adults. No juveniles of V. ardabilia have been found.
POPULATION GENETICS AND SPECIATION
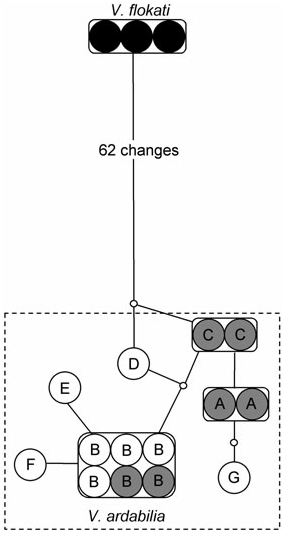
The resulting COI dataset consists of 585 characters of which 318 are variable, and 277 are parsimonyinformative. The Kimura 2 Parameter (K2P) nucleotide distance between V. flokati and V. ardabilia is 11.7%. Of the 63 substitutions separating V. flokati from V. ardabilia in the studied COI fragment, one occurs in a second position and six in first positions. Of these substitutions, five result in amino acid changes. The parsimony network constructed with TCS using a 95% connection limit separated the two species into two networks. The three sequenced V. flokati specimens all had identical COI-haplotypes, whereas seven different haplotypes were observed in the 14 sequenced V. ardabilia specimens from Norway and Sweden ( Fig. 5 View Figure 5 ). Within V. ardabilia , the most common haplotype group (B) was shared by six individuals, with the remaining haplotypes present in either one or two individuals ( Fig. 5 View Figure 5 ). Two haplotype groups (A, C) were present only in individuals collected from the Norwegian fish farm, but the most common haplotype group (B) was present in individuals from both localities, implying gene-flow between the Norwegian and Swedish populations.
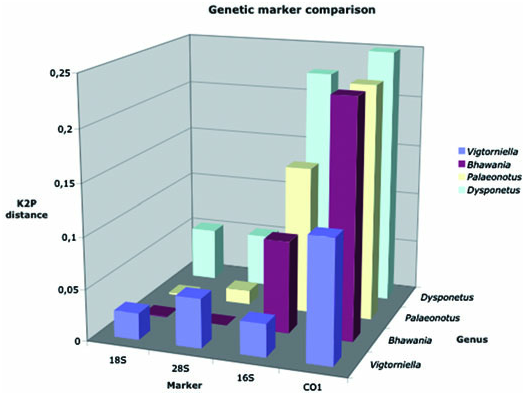
A comparison was made of species pair divergence (K2P distances) for four genetic markers (18S, 28S, 16S, and COI) within four congeneric species pairs within Chrysopetalidae ( Fig. 6 View Figure 6 ). Within Bhawania, Palaeonotus , and Dysponetus , the congeneric pairs show distinct morphological differences, whereas within Vigtorniella , the speciation is morphologically cryptic. This cryptic speciation process in a high sulphide habitat has a different distribution of genetic divergence. Whilst the other species pairs follow the
CRYPTIC SPECIATION IN ORGANIC-RICH HABITATS 781
convention of much greater divergence in mitochondrial markers (16S and COI) compared with nuclear markers (18S and 28S), Vigtorniella is different. These congeners show much higher nuclear gene distances relative to the other species pairs, with higher nuclear 28S divergence than mitochondrial 16S divergence.
PHYLOGENETIC POSITION
The combined dataset consists of 2917 characters, 810 variable, of which 604 are parsimony-informative. With high levels of support, all our analyses suggest that V. ardabilia is a sister taxon to V. flokati ( Fig. 7A, B View Figure 7 ). The tree topology from the BA and ML analyses differed only in the position of Chrysopetalum debile , which in the ML analysis was sister to Bhawania , although with low bootstrap support (not shown). In the BA, Chrysopetalum was instead sister to Paleanotus ( Fig. 7A View Figure 7 ). Both analyses suggest that the genera Dysponetus and Vigtorniella are sister taxa, and form a sister group to other chrysopetalid taxa ( Fig. 7A View Figure 7 ). Also in the PA, Dysponetus and Vigtorniella were sister taxa, while the relationships of the other worms in the Chrysopetalidae group were unresolved ( Fig. 7B View Figure 7 ).
ECOLOGY AND LIFE HISTORY
The ability to culture V. ardabilia on aquariummaintained whale bones has allowed us to make some further observations on feeding and reproductive biology. Live observations indicated that the worms move actively over the surface of the whale bones, sometimes crawling inside holes in the bone matrix. We found the worms most frequently associated with white bacterial mat, composed of several undescribed species of giant filamentous sulphide-eating bacteria, including Beggiatoa (Hans Roey, pers. comm.). The worms were found on the same bones as Osedax mucofloris Glover et al., 2005a and several undescribed new species of Ophryotrocha Claparède & Mecznikow, 1869 (H. Wiklund, unpubl. data). Vigtorniella ardabilia was observed actively feeding on white bacterial mat, using its pharynx to package the mat into gut pellets. On bones that had been heavily infested with Osedax , V. ardabilia was able to move inside the soft, burrowed bone tissue, possibly to avoid predators.
Gonochorism was observed, with females containing eggs from segment 6 through to the posterior end of the body and males with spermatids in segments 6–8. Eggs were 40–50 Mm in size. Of 15 specimens measured, size ranged from 5 to 35 mm and the number of segments from 28 to 82, with no statistical difference between the size of female vs. male specimens. Eggs were observed in eight of the 15 specimens examined. A comparison with V. flokati in size and number of segments showed that there is no size difference between the two species (Fig. 8).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |