
Poteranthera inopinata Almeda & R.B.Pacifico, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.613.3.6 |
|
persistent identifier |
https://treatment.plazi.org/id/03E98781-FFCE-FFE7-FF2B-8650FA24FE30 |
|
treatment provided by |
Plazi |
|
scientific name |
Poteranthera inopinata Almeda & R.B.Pacifico |
| status |
sp. nov. |
Poteranthera inopinata Almeda & R.B.Pacifico View in CoL , sp. nov. ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 4A View FIGURE 4 )
Type:— BOLIVIA. Santa Cruz: Velasco Province. Parque Nacional Noel Kempff Mercado, Pampa Grande de Bella Vista , 13°42’10.7”S, 61°31’58.1”W, 180 m, 11 August 1995 (fl., fr.), R. Guillén et al. 3903 (holotype US-02920817 !; isotypes: CAS-0437832!, F-2248625, digital image!, MO-979360 , NY-02707947!, USZ, n.v.) GoogleMaps .
Diagnosis:— Poteranthera inopinata can be distinguished from P. warmingii by its uppermost branchlet nodes copiously covered with gland-tipped trichomes (vs. nodes that are glabrous or only beset with three or fewer trichomes), shorter petiolate (0.5–2 mm long) leaf blades that are 3–10 mm long (vs. sessile leaf blades that are 15–20 mm long), conspicuously serrate blade margins (vs. subentire to inconspicuously serrulate margins), and petals tipped with a glandular trichome 0.5–0.75 mm long (vs. petals tipped with an eglandular trichome 0.25–0.5 mm long).
Erect openly branched annual herbs 10–55 cm tall with slender elongate branches. Upper cauline internodes 0.5–3 cm long, often reddish, quadrangular and moderately covered with spreading gland-tipped trichomes mostly 0.5–1 mm long, the angles narrowly winged (0.25–0.5 mm wide), the nodes densely covered with spreading gland-tipped trichomes 0.5–1.5 mm long. Leaves of a pair equal in size, chartaceous, subsessile to distinctly petiolate, erect to ascending (when dry), ± equal to or greatly exceeded in length by the internodes, petioles 0.5–2 mm long, flattened to canaliculate on the adaxial surface and covered with trichomes like the internodes; blades 3–10 × 3–7 mm, ovate to ovate-elliptic varying to elliptic, brown when dry, base commonly rounded varying to obtuse or attenuate (on uppermost leaves), apex acute, terminating in a gland-tipped trichome ca. 1 mm long, margins conspicuously serrate with each serration terminating in a gland-tipped trichome ca. 1 mm long, 3–5-nerved from the base, midvein evident for a portion of its length on the adaxial surface, prominent and elevated on the abaxial surface, secondary veins evident for the entire length of the blade on the abaxial surface but not evident on the adaxial surface, tertiary veins not evident on either surface, both surfaces moderately to sparsely covered with spreading gland-tipped trichomes 0.5–1.25 mm long. Flowers 5-merous, diplostemonous, initially terminal but becoming axillary and solitary on upper branches with shoot elongation, ebracteolate, pedicels 0.5–1.5 mm long (lengthening up to 3.5 mm on fruiting hypanthia), mostly sparsely beset with a mixture of inconspicuous sessile deciduous glands and spreading persistent gland-tipped trichomes like those of the internodes; hypanthia (at anthesis) 2.5–3 × 2–2.5 mm, widest at the middle, campanulate but becoming distinctly urceolate and constricted distally below the calyx lobes in fruit, sparsely covered with spreading gland-tipped trichomes 0.5–1 mm long, calyx tube ca. 0.25 mm long; the persistent calyx lobes 2.5–3.5 mm long and 1 mm wide at the base, narrowly triangular, apex attenuate, terminating in an apical glandular trichome 1–2 mm long, adaxial surface glabrous, abaxial surface and margins beset with gland-tipped trichomes like the hypanthia; petals 4.4–7.0 × 2.7–4.8 mm, pink, obovate, apex apiculate and bearing a gland-tipped trichome 0.5–0.75 mm long, margins entire and bearing minute inconspicuous deciduous glandular trichomes, otherwise glabrous on both surfaces; stamens 10, dimetric and dimorphic: antesepalous (larger) stamens 5, filaments 2.6–4 mm long, pedoconnectives 2.8–4 mm long, appendages 0.3–0.5 mm long, yellow, blunt and ± rounded and retuse, thecae 2–3 mm long, oblong-subulate, pinkish-brown (when dry), the surface smooth (tetrasporangiate), rostrum 0.4–0.5 mm long, white, the circular pore ca. 0.18–0.35 mm wide; antepetalous (smaller) stamens 5, filaments 2.6–3.5 mm long, pedoconnectives 0.9–1 mm long, appendages 0.2–0.25 mm long, yellow, blunt, rounded and retuse, thecae 1.5–2.5 mm long, oblong-subulate, pink-brown (when dry), surface smooth (tetrasporangiate), rostrum ca. 0.25–0.35 mm wide, white, the circular pore ca. 0.13–0.25 mm wide. Ovary (at anthesis) 1.4–3 × 1–2.5 mm, ± oblong, superior, glabrous, 3-locular; styles 6–8 mm long, glabrous, ± straight or somewhat declinate, stigmas punctiform. Fruit a loculicidal capsule 3–3.5 × 3–3.5 mm, brown, globose, 3-valvate, the enveloping hypanthia and calyx lobes rupturing and flaking away with age, dehiscent from apex to the base (basipetal), columella deciduous. Seeds 0.36–0.45 × 0.64–0.75 mm, tan, oblong-reniform, testa shallowly foveolate, raphal zone nearly circular, ca. 20 % the length of the seed.
Paratypes:― BOLIVIA. Santa Cruz: Velasco Province. Parque Nacional Noel Kempff Mercado, Los Fierros , 14°37'15"S, 60°51'10"W, 250 m, 4 May 1994 (fl.), E. Gutiérrrez et al. 736 ( USZ); Parque Nacional Noel Kempff Mercado , Campamento Flor de Oro , 13°32'45"S, 61°01'07"W, 200 m, 27 May 1994 (fl., fr.), E. Gutiérrez et al. 1306 ( MO, US-02924253 !); Campo Grande Guarasug’we plano, 13°43'47.7"S, 61°27'49.8"W, ca. 230 m, 14 August 1995 (fl., fr.), S. Halloy et al. 4431 (NY-02710501!), S. Halloy et al. 4432 (NY-02707950!), S. Halloy et al. 4433 (NY-0270950!) GoogleMaps .
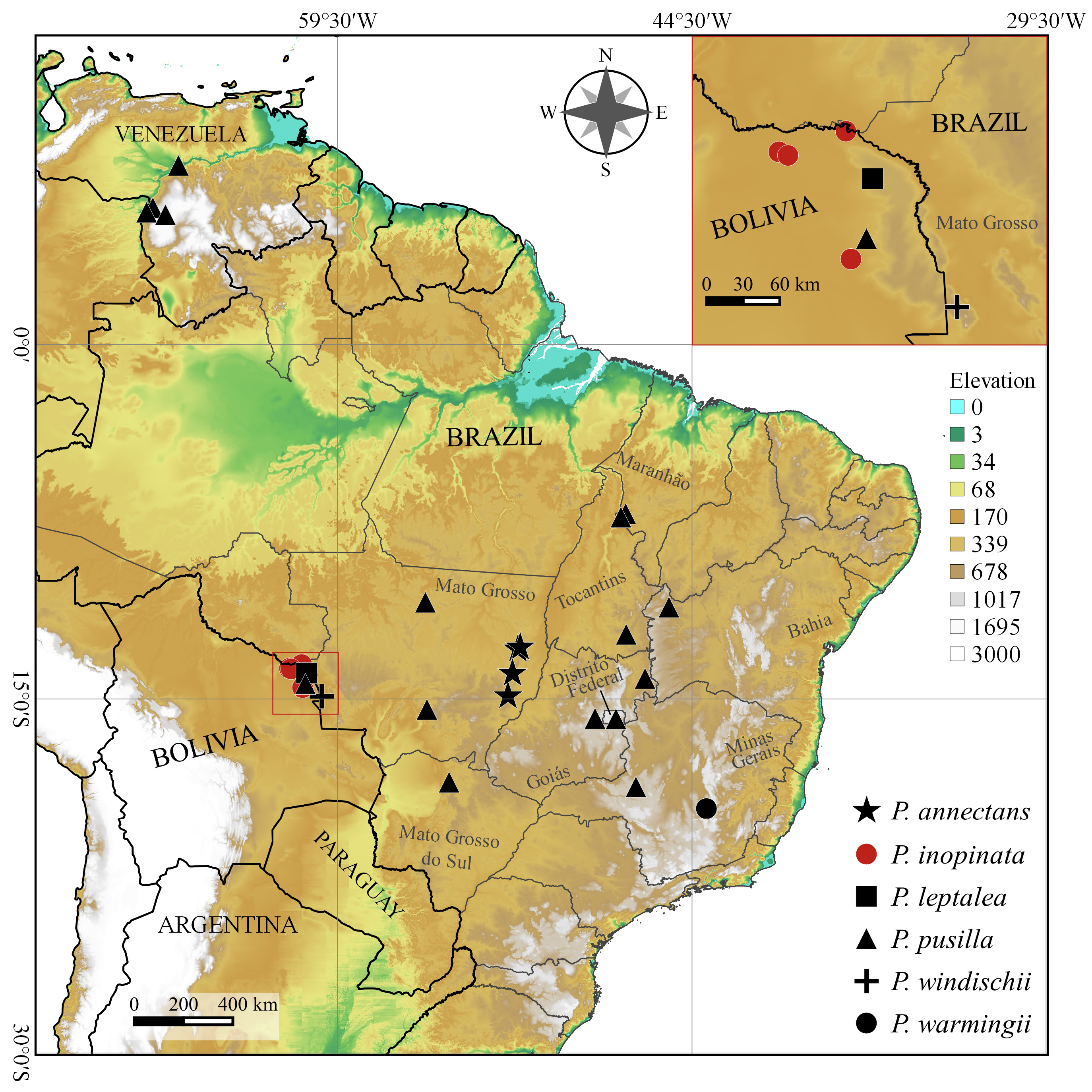
Distribution, Habitat, and Phenology:— Poteranthera inopinata is known only from Noel Kempff Mercado National Park in eastern Bolivia at an elevation of 180–250 m ( Fig. 3 View FIGURE 3 ). Judging from known collections it appears to be local and uncommon in the pampas húmido (wet grasslands) and temporarily flooded pampas with termitaria. Because of its ephemeral annual life cycle it has likely been overlooked. It has been collected in flower and fruit in May and in August. The most recent collections were made in 1995.
Conservation Status:—Both EOO (555.541 km 2) and AOO (12.000 km 2) calculations estimated with GeoCAT support a conservation assessment of Endangered (EN): B2ab(iii) for Poteranthera inopinata . All but one of the known collections come from the northernmost part of Noel Kempff Mercado National Park ( Fig. 3 View FIGURE 3 ). Because of its annual life cycle, habitat specificity, and few known collections, it is difficult to gauge how common this species is within the park boundaries. Thus, all of these factors make it potentially vulnerable to decline over time.
Based on IUCN’s World Heritage Outlook Assessment (2020), the protected status of Noel Kempff Mercado National Park is described as “good with some concerns”. Its conservation status continues to benefit from its immense size and isolation from areas of human settlement. Wildfires are an ongoing reason for concern because they cannot be managed effectively. Like many parks in developing countries, Noel Kempff Mercado NP is short-staffed and underfunded. In addition to the ongoing threat of wildfires, the park suffers from the entry of illegal settlers, narcotics trafficking, illegal grazing of livestock, hunting, commercial fishing, and illegal logging ( IUCN 2022). Nothing is yet known about the future effects of climate change.
Etymology: — The specific epithet is derived from the Latin word, inopinatus, which means unexpected, unforeseen, or surprising. It highlights the fact that this species, which superficially resembles and had been identified as Acisanthera uniflora ( Vahl 1791: 48) Gleason (1950: 346) , was overlooked, so its discovery was surprising and unexpected.
Affinities: — The few collections of Poteranthera inopinata have been variously identified as Acisanthera uniflora , Microlicia D. Don (1823: 301) , or Siphanthera Pohl ex Candolle (1828: 121) . Our new species differs from Acisanthera uniflora , which occurs in Bolivia but apparently not in Noel Kempff M. National Park ( Peña et al. 77 & 241, both at F), in having conspicuously serrate leaf blades with each serration bearing a long gland-tipped trichome (vs. minutely ciliolate-denticulate along entire margins), narrowly triangular calyx lobes (vs. linear-oblong calyx lobes), shallowly retuse staminal appendages (vs. deeply lobed appendages) and seeds that are oblong-reniform ( Fig. 2B & 2C View FIGURE 2 ) with a shallow foveolate testa ( Fig. 2D View FIGURE 2 ) (vs. seeds that are cochleate and often sometimes costate with a deeply foveolate testa) (see Wurdack et al. 1993: 27 for a diagnostic illustration). All species of Poteranthera differ from Microlicia in their annual herbaceous habit (vs. perennial woody habit), and consistent presence of only long gland-tipped trichomes on vegetative and floral parts (vs. a wide array and commonly a mixture of indument types on vegetative and floral parts), and hypanthia that are conspicuously constricted at the apex (vs. hypanthia that are typically not constricted at the apex) (see Versiane et al. 2021). If hypanthia are constricted, as they are in a few species of the Lavoisiera clade of Microlicia , then the ovary is always at least partially inferior and the number of locules ranges from five to eight (see Martins & Almeda 2017). One other species, Microlicia viminalis ( Candolle 1828: 103) Triana (1871: 28) , has hypanthia that are conspicuously constricted distally at maturity but it is readily separated from all species of Poteranthera by its woody habit, well-developed xylopodium, coriaceous sessile leaves (3–9-nerved) that are pungent apically and have entire margins, and larger seeds (1–2.5 mm long) (see Diniz & Silva 2020). All Poteranthera species differ from Siphanthera in having stout gland-tipped trichomes on the leaf margins and oblong-reniform seeds with a foveolate testa (vs. stout gland-tipped trichomes absent and seeds that are ellipsoid to lacrimiform or subreniform with a testa of areolate cells that are elongate-tetragonal to elongate polygonal and gradually smaller toward both ends) ( Almeda & Robinson 2011). Besides, all species of Poteranthera , except P. leptalea , have 5-merous flowers with 3- locular ovaries, whereas all species of Siphanthera are consistently 4-merous and have 2-locular ovaries.
Among described species of Poteranthera , only P. warmingii and P. inopinata share a 5-merous flower, a 3- locular ovary, 3–5-nerved leaf blades, and both cycles of stamens that are fertile. The salient characters that distinguish these two species are enumerated in the diagnosis. These two species are also allopatric, with the former known only from the holotype which was collected in the Lago Santa region of Minas Gerais, Brazil ( Almeda & Pacifico 2018).
Poteranthera inopinata differs from all of its congeners in having a combination of narrowly winged upper cauline internodes, ovate to ovate-elliptic leaves that have conspicuously serrate margins with each serration tipped by a glandular trichome 1 mm long ( Fig. 1 View FIGURE 1 ), petals that are minutely and deciduously glandular on the margins and tipped with a persistent glandular trichome (0.5–0.75 mm) at the apiculate apex, and ten stamens that are dimetric and dimorphic. It can be distinguished by the diagnostic characters noted in the following key which is modified from Almeda & Pacifico (2018) and Kriebel (2012).
Notes:—There are conflicting reports in the literature regarding the production of staminodia in flowers of Poteranthera pusilla ( Bongard 1838: 137) . In the protologue (see pl. 8, fig. 1) the dissected flower of this species shows no staminodia. However, Cogniaux (1883) describes the species as having “stamina sterilia dimidio minora”, but the figure of a dissected flower in the same publication (pl. 47, fig. IV) shows no staminodia. Wurdack (1963) also states that staminodia are absent in this species. In Kriebel’s (2012) key to the three species of Poteranthera that were known at the time he states that P. pusilla lacks staminodia altogether but describes them as “staminodes with filaments 0.3–0.5 mm long, the thecae ca. 0.1–0.2 mm ....” in the species description. Versiane et al. (2016) also report staminodia for this species. In other genera with haplostemonous species such as Siphanthera , staminodia are produced in some but not necessarily all populations of a species ( Almeda & Robinson 2011). Similar variation in the production of staminodia may occur in the geographically widespread P. pusilla ( Fig. 3 View FIGURE 3 ). The key below reflects this possibility pending more information and critical field observations. We encourage collectors to make special note of this feature when this species is encountered in the field.
At US there is another specimen of R. Guillén et al. 3903 (US-02924448). It consists of two branches of Siphanthera subtilis Pohl ex Candolle (1828: 121) that were evidently mixed with the type of P. inopinata but separated and mounted on a separate sheet with the same duplicated label. This sheet ( Fig. 4B View FIGURE 4 ) has a separate barcode and is not part of the type collection of P. inopinata .
Key to the species of Poteranthera
1. Flowers 4-merous, ovary 2-locular...................................................................................................................................... P. leptalea
1. Flowers 5-merous, ovary 3-locular.
2. Leaf blades 3–5-nerved, ovate to ovate-elliptic or ovate-lanceolate; hypanthia sparsely to moderately covered with spreading gland-tipped trichomes; flowers with both cycles of stamens well-developed and fertile.
3. Leaf blades 15–20 × 3–6.5 mm, sessile, ovate-lanceolate, margins subentire to remotely and inconspicuously serrulate with glandular trichomes 0.5–1.3 mm long; uppermost nodes glabrous or with three or fewer gland-tipped trichomes 0.5 mm long; petals entire and tipped with a single eglandular trichome 0.25–0.5 mm long............................................................... P. warmingii
3. Leaf blades 3–10 × 3–7 mm on petioles 0.5–2 mm long, ovate to ovate-elliptic, margins conspicuously serrate with each serration tipped by a glandular trichome 1 mm long; uppermost nodes covered with numerous spreading gland-tipped trichomes 0.5–1.5 mm long; petals entire but the margins inconspicuously beset with minute gland-tipped trichomes and tipped with a solitary trichome 0.5–1 mm long.................................................................................................................................................................. P. inopinata
2. Leaf blades 1-nerved, linear to oblong; hypanthia glabrous; flowers with one cycle of stamens reduced to staminodia or absent altogether.
4. Flowers haplostemonous or diplostemonous; stamens 5 or if 10 (then 5 are staminodial); anther thecae ovate with a wide truncate ventrally inclined pore; the larger (antesepalous) cycle of stamens with a pedoconnective 0.1–0.3 mm long..................... P. pusilla
4. Flowers diplostemonous, stamens 10 (including 5 staminodia); anther thecae subulate with a cucullate pore, the larger (antesepalous) cycle of stamens with a pedoconnective 1–5 mm long; smaller (antepetalous) cycle of stamens with a pedoconnective 1–5 mm long.
5. Cauline internodes glabrous; petals 5 × 3 mm, the apex acute, bearing a glandular trichome 0.4–1 mm long; anther thecae of large stamens yellow, pedoconnectives of the larger (antesepalous) stamens prolonged to 1 mm .......................................... P. annectans
5. Cauline internodes sparsely covered with gland-tipped trichomes 0.25–1 mm long; petals 7–9 × 6–7 mm, the apex rounded, lacking a gland-tipped trichome; anther thecae of large stamens purple, pedoconnectives of the larger (antesepalous) stamens prolonged 3.8–5 mm ........................................................................................................................................................ P. windischii



| USZ |
Museo de Historia Natural Noel Kempff Mercado -- Universidad Autónoma Gabriel René Moreno |
| MO |
Missouri Botanical Garden |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |