
Acanthechiniscus goedeni ( Grigarick, Mihelčič & Schuster, 1964 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5227.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:8A15E9F9-894D-4C37-A41D-A066AEE3E07C |
|
DOI |
https://doi.org/10.5281/zenodo.7525214 |
|
persistent identifier |
https://treatment.plazi.org/id/03E4ED39-CB4E-FFBB-FF42-34A7FC1BF873 |
|
treatment provided by |
Plazi |
|
scientific name |
Acanthechiniscus goedeni ( Grigarick, Mihelčič & Schuster, 1964 ) |
| status |
|
Acanthechiniscus goedeni ( Grigarick, Mihelčič & Schuster, 1964) View in CoL
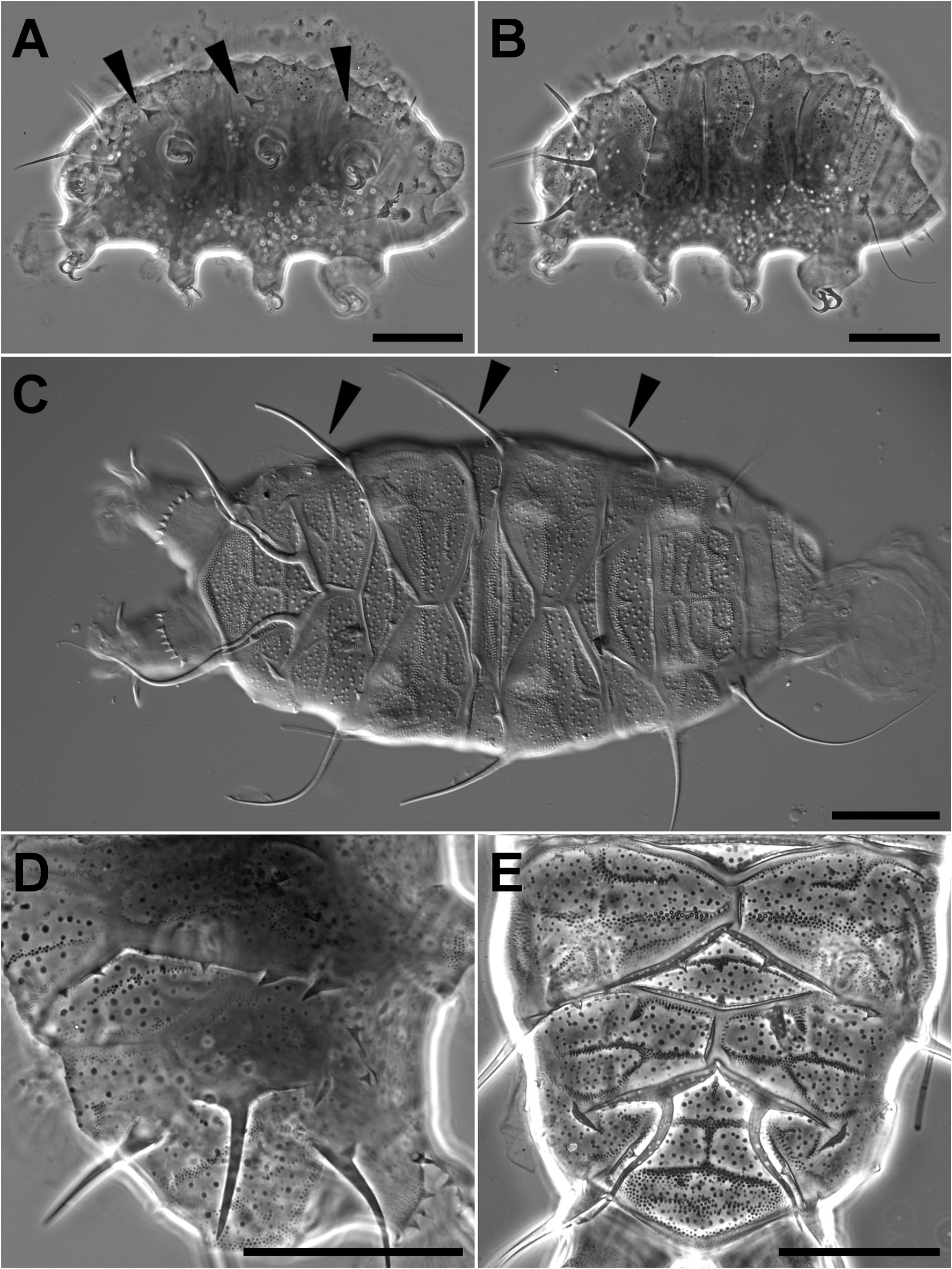
( Figure1 View FIGURE 1 )
Material examined and repository
Voucher specimens are deposited in the Invertebrate Zoology Collections, Royal BC Museum ( RBCM), 675 Belleville Street , Victoria, BC Canada (sample code: S198 HC 2018-14-1), RBCM022-0030 View Materials - 001 View Materials ; (sample code: S1904 GSB-1c), RBCM022-00031 View Materials - 1 View Materials ; and (sample code: S1913 GSB-7), RBCM022-0032 View Materials - 001 View Materials ).
Description of the larva. Length 230μm, body plump ( Fig. 1A, B View FIGURE 1 ). Eyes not visible after mounting. On the head, a full set of appendages is present: cirrus internus (8.5 μm), cephalic papilla (7.0 μm), cirrus externus (10.8 μm). Dorsal plates as described by Grigarick, Mihelčič & Schuster (1964) and Kathman & Dastych (1990) ( Fig. 1 B, C View FIGURE 1 ), but with fewer and less dense granules than the adult ( Fig. 1D, E View FIGURE 1 ). Cirrus A present (64.4 μm) with clava (5.0 μm). Very small and stout lateral spines in B (7.3 μm), C (9.1 μm), and D (6.8 μm) (in the same positions that would be filaments in the adult) ( Fig. 1A View FIGURE 1 ). Small dorsolateral spinules are present above the spines in B, C, and D. In D, a pair of additional dorsal spinules are present, which are smaller than the ones in dorsolateral position D. On the pseudosegmental plate, two long spines ( Fig. 1 B, D View FIGURE 1 ) (29.7 μm) are present in the dorsal position (similarly to the adult, Fig. 1 C, E View FIGURE 1 ), in addition to small dorsolateral small spinules and two spines in the lateral position. Two long spines (34.1 μm) on the caudal plate E are present. Leg I with small spine (1.8 μm), leg IV with papilla (5.3 μm) and dentate collar with 6 teeth present. Two claws present, both with spur oriented downward starting at about 30% of the claw length on each leg ( Fig. 1B View FIGURE 1 ) (adult with four claws and spur on the two internal claws). Lengths of claws from I to IV are: 11.6 μm, 9.5 μm, 10.2 μm, and 11.3 μm, respectively. No anus or gonopore observed.
DNA. Four sequences were obtained from one individual:
• SSU: GenBank OP730698 View Materials
• LSU: GenBank OP730697 View Materials
• COI: GenBank OP729912 View Materials
• ITS2: GenBank OP734819 View Materials
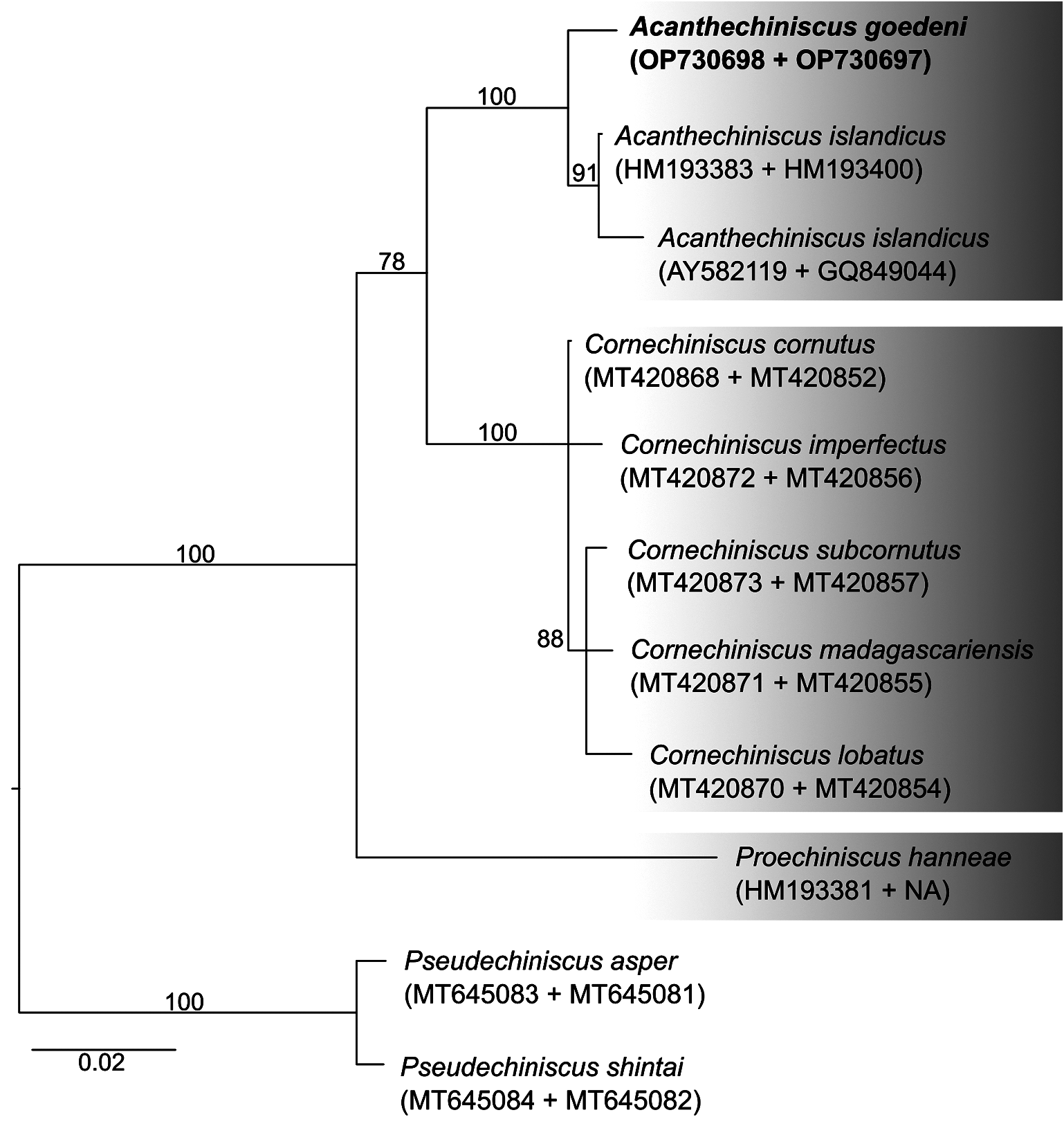
Phylogenetic position. The phylogenetic reconstruction ( Figure 2 View FIGURE 2 ) recovered A. goedeni as closely related to A. islandicus . The relatedness of A. goedeni to A. islandicus had already been indicated by Grigarick et al. (1964) based on some similarities in the chaetotaxy and in the plate ornamentation. The phylogenetic reconstruction supports the generic attribution of A. goedeni , as proposed by Vecchi et al. (2016) and Gąsiorek & Michalczyk (2020).
The discovery of a new population of A. goedeni expands the range of this species to the north in British Columbia ( Canada) by about 250 km, which is consistent with all the previous records that point to this species as being endemic to the north-west coast of North America. As pointed out by Bertolani et al. (1984), the ontogeny of echiniscid lateral appendages is variable between species and genera. In another Acanthechiniscus species ( A. victor , synonym A. tridentifer ), Bertolani et al. (1984) reported that the number of trunk appendages remained the same, but they increased in length during development. An identical developmental pattern was also found in A. islandicus (Piotr Gąsiorek, pers. comm.).
A similar general pattern (increase in the number and size of trunk appendages during development) has been reported from different echiniscid taxa (see for example species in the genus Echiniscus ; Claxton, 1996; Gąsiorek et al., 2022). Other developmental pathways found in echiniscids, but not observed in this study of A. goedeni , include the blurred distinction between dorsal plates (in Proechiniscus and Pseudechiniscus ; Kristensen, 1987; Gąsiorek et a., 2022) and a simultaneous increase in size in lateral trunk appendages with a reduction in both size and number of dorsal trunk appendages (in Mopsechiniscus ; Dastych, 2001).
We are grateful to the Hakai Institute, Tula Foundation- Eric Peterson and Christina Munck for making research on Calvert Island possible. We also wish to thank Gustav Paulay (University of Florida), Matt Lemay (Hakai), and Matt Whalen (UBC-Hakai) for facilitating collecting opportunities, Gillian Sadlier-Brown (Hakai) for collecting part of the samples used in this study, and Hugh MacIntosh (Royal BC Museum) for assistance with the processing of field samples. We are grateful to Daniel Stec (Institute of Systematics and Evolution of Animals, Polish Academy of Sciences) for acquiring microphotographs of the specimens belonging to the population examined in this study. We also wish to thank Piotr Gąsiorek and an anonymous reviewer for their constructive comments and suggestions. This study was supported by an Academy of Finland Fellowships to S.C. (#314219 and #335759).
| RBCM |
Royal British Columbia Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |