
Syllis broomensis Hartmann
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4052.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:52083710-01B3-4CF1-A6CE-5A09419F7D25 |
|
DOI |
https://doi.org/10.5281/zenodo.6112218 |
|
persistent identifier |
https://treatment.plazi.org/id/03E4CD0F-FFC2-FFCB-9199-FA66FED28379 |
|
treatment provided by |
Plazi |
|
scientific name |
Syllis broomensis Hartmann |
| status |
|
Syllis broomensis Hartmann View in CoL – Schröder, 1979 n. comb.
Figures 1 View FIGURE 1 , 2 View FIGURE 2
Typosyllis (Langerhansia) broomensis Hartmann-Schröder, 1979: 88 View in CoL , Figs 50–56; 1984: 18; 1989: 23; 1990: 48; 1991: 32. Typosyllis broomensis View in CoL . Licher 1999: 70, Fig. 32.
Typosyllis (Langerhansia) cervantensis Hartmann-Schröder, 1981 View in CoL , through synonymy by Licher 1999: 70.
Material examined. Type specimens. Typosyllis (Langerhansia) broomensis . WESTERN AUSTRALIA. Broome, fine sand with detritus, intertidal, 8 Sept 1975, coll. Hartmann-Schröder, 2 paratypes ( HZM P- 15465).
Non-type specimens. AUSTRALIA. QUEENSLAND. Lizard Island, High Rock, 14°49'34"S, 145°33'08"E, sand, 20 m depth, 11 Sept 2010, CReefs Lizard Island Expedition, 1 specimen (AM W41705). Calliope River, 23°49'S, 151°13'E, 1974, 1 specimen mounted for SEM (AM W42547.001). WESTERN AUSTRALIA. Cassini Island, 13°57'22"S, 125°37'23"E, fore-reef slope, coral rubble, 11 m depth, 15 Oct 2010, Woodside Kimberley Survey 2010, 1 specimen (AM W42539). Southeast end of Long Island, Goss Passage, 28°28'48"S, 113°46'30"E, dead coral embedded in calcareous substrate, 30 m depth, 22 May 1994, 1 specimen (AM W46277). Exmouth Gulf, beach at north end of Bundegi Reef, 21°49'S, 114°11'E, rocky rubble, coralline algae with green epiphyte, 1– 2 m depth, 4 Jan 1984, 2 specimens (AM W46281).
PHILIPPINES. LUZON ISLAND. “Sepok Wall”, between Balayan Bay and Batangas Bay, 13º41'02"N, 120º53'45"E, coral rubble, 6–13 m depth, 10 Dec 2010, 1 specimen ( MNCN 16.01/16868). NEW ZEALAND. Kermadec Islands, Raoul Island, Kermadec Biodiscovery Expedition, 2011: “Fishing Rock” landing, 29°15'03"S, 177°54'12"W, under boulders in rockpools, algal turf, 1 m depth, 18 May 2011, 5 specimens (AM W42559); “Fishing Rock” 29°15'03"S, 177°54'12"W, under boulders in rockpools, some loose rocks and coral cover in pool, 1 m depth, 18 May 2011, 5 specimens (AM W42560; South side of Te Konui Point, 29°18'32"S, 177°53'45"W, base of boulders with coarse sand and gravel, tufting algae, 21 m depth, 14 May 2011, 2 specimens (AM W42561).
Morphologically similar species. Syllis garciai ( Campoy, 1982) . SPAIN. Almería, Playa de los Genoveses, Cabo de Gata, 4 m depth, Nov 1986, 1 specimen ( MNCN 16.01/6998). Syllis yallingupensis ( Hartmann-Schröder, 1982) , 9 Jul 1993, BANGAWW NT316, 2 specimens (AM W29546).
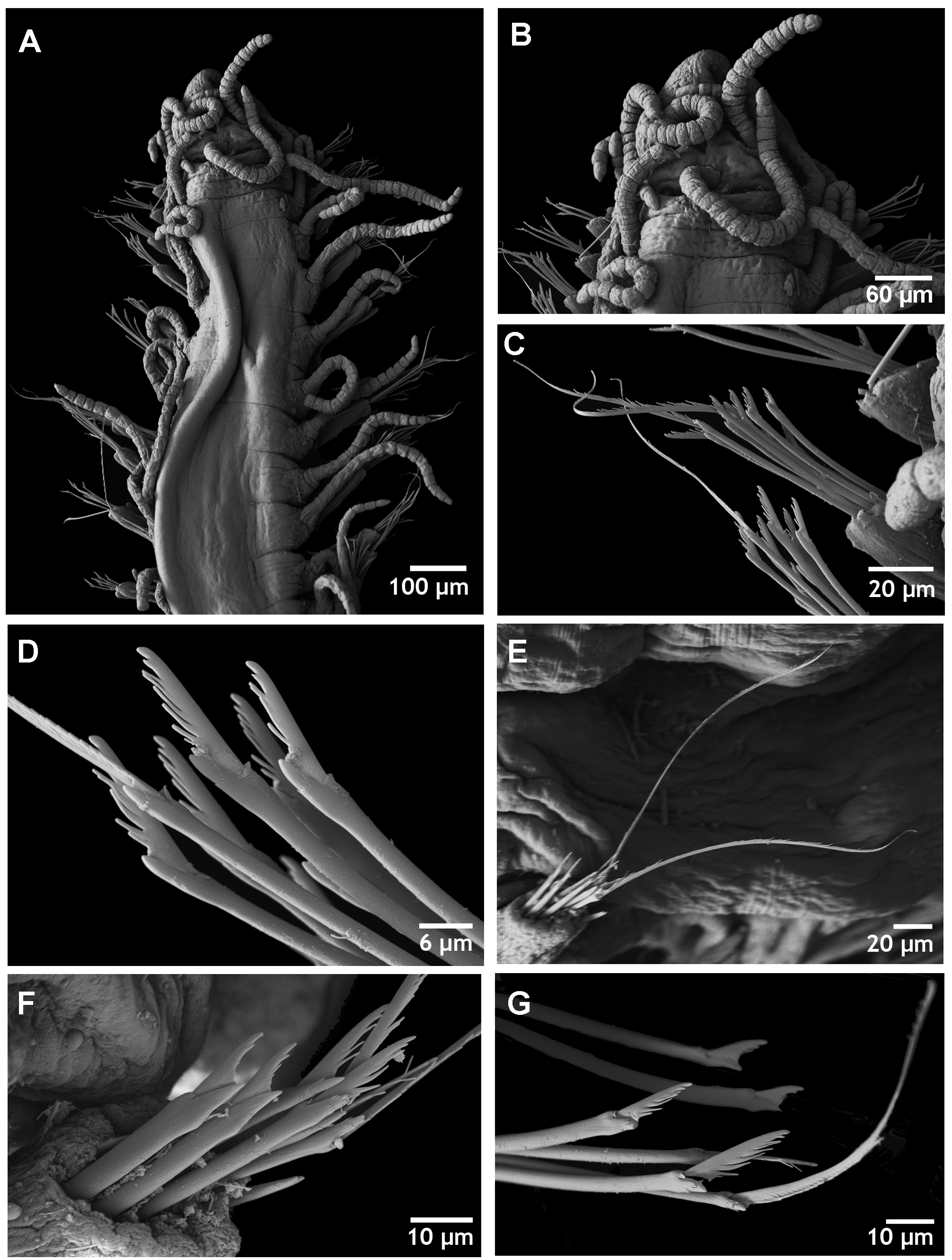
Description. Longest complete specimen examined 5 mm long, 0.25 mm wide, with 50 segments. Slender elongated body, lacking colour patterns. Oval prostomium, with two pairs of red eyes in open trapezoidal arrangement, anterior ones slightly larger than posterior ones ( Fig. 1 View FIGURE 1 A), and 2 anterior eyespots. Median antenna inserted near posterior margin of prostomium, between posterior eyes, longer than the combined length of prostomium and palps, with 19–23 articles ( Figs 1 View FIGURE 1 A, 2A–B); lateral antennae distinctly shorter, inserted close to anterior margin of prostomium, with 17–19 articles ( Figs 1 View FIGURE 1 A, 2A–B). Triangular palps, longer than prostomium, fused at base, with distinct median groove. Nuchal organs forming a ciliary groove between prostomium and peristomium ( Fig. 2 View FIGURE 2 B). Peristomium slightly shorter than subsequent segments. Dorsal tentacular cirri longer than antennae, with 24–26 articles, ventral ones shorter, with 10–12 articles ( Figs 1 View FIGURE 1 A, 2A). Dorsal cirri of anteriormost segments with 22–24 articles. Dorsal cirri of midbody and posterior chaetigers alternating long (11–21 articles) and short (9–11 articles) ( Figs 1 View FIGURE 1 A, 2A). Digitiform, elongated ventral cirri, inserted proximally and reaching distal end of parapodia. Anterior and midbody parapodia with about nine heterogomph falciger chaetae, posterior parapodia with four to five chaetae. Parapodia throughout with one to three dorsal pseudospiniger chaetae, unidentate, with long and thin spines on margin; blades about 84 µm long on anterior and midbody parapodia and 104 µm long on posterior ones ( Figs 1 View FIGURE 1 C, E, 2C, E, G). Remaining compound chaetae shorter, with bidentate blades, both teeth similar, and long, fine, slightly curved spines on margin, reaching level of proximal tooth; blades and shafts of posterior chaetae shorter and wider than those from anterior and midbody parapodia, with shorter spines on cutting edge ( Figs 1 View FIGURE 1 D, F, 2D, F, G). Thick, smooth, distally bidentate dorsal simple chaetae on posterior parapodia only ( Fig. 1 View FIGURE 1 I). Ventral simple chaetae not seen. Anterior parapodia each with 4–5 aciculae, distally straight or slightly curved ( Fig. 1 View FIGURE 1 G); midbody parapodia with two aciculae each, one straight, slightly curved and other acuminate, distally pointed ( Fig. 1 View FIGURE 1 H); each posterior parapodium with single, acuminate, distally pointed acicula ( Fig. 1 View FIGURE 1 J). Pharynx through 6–8 segments; pharyngeal tooth on anterior margin ( Fig. 1 View FIGURE 1 A). Proventricle extending through 7–8 segments, with about 30 rows of muscle cells ( Fig. 1 View FIGURE 1 A). Pygidium conical with two short anal cirri and median stylus.
Remarks. The AM specimens seemed to be juveniles, but their morphology agree with the examined paratypes (HZM P-15465) and with Licher’s (1999) re-description, except for the smaller size, the pseudospiniger chaetae that are slightly shorter on anterior parapodia, and the smaller number of aciculae per parapodium; the proventricle of one of the paratypes was also larger (through 10 segments) than those of our specimens. The specimens collected in the Philippines also agreed with the type material, except for the length of the dorsal cirri being shorter in the former, with 17–18 articles. All the differences found in our specimens could indicate intraspecific variability depending on the ontogenetic stage of the specimens, or it could also mean that they are different, geographically distant species. The most similar species in terms of morphology to S. broomensis n. comb. is the Mediterranean S. garciai ( Campoy, 1982) and the Australian S. yallingupensis ( Hartmann-Schröder, 1982) . The former only differs in the presence of a ventral simple chaetae on posterior parapodia and the latter in the length of dorsal blades (shorter than those from S. broomensis n. comb.), the number and shape of aciculate (see Licher 1999 for comparison) and the absence of dorsal simple chaetae in posterior parapodia.
Habitat. Mainly in coral rubble, coarse sand and algae.
Distribution. Australia (Queensland, Western Australia, New South Wales), the Philippines, New Zealand (first record).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Syllis broomensis Hartmann
| Álvarez-Campos, Patricia, Riesgo, Ana, Hutchings, Pat & Martín, Guillermo San 2015 |
Typosyllis (Langerhansia) cervantensis Hartmann-Schröder, 1981
| Licher 1999: 70 |
Typosyllis (Langerhansia) broomensis Hartmann-Schröder, 1979 : 88
| Licher 1999: 70 |
| Hartmann-Schroder 1979: 88 |