
Hermippus gavi, Sankaran, Pradeep M., Jobi, Malamel J., Joseph, Mathew M. & Sebastian, Pothalil A., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3893.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:BA1E5A6D-FB15-47BD-BDFA-F661689CAC2C |
|
DOI |
https://doi.org/10.5281/zenodo.6144827 |
|
persistent identifier |
https://treatment.plazi.org/id/03E0522E-FFC9-2504-22F9-FACAFB03FC7C |
|
treatment provided by |
Plazi |
|
scientific name |
Hermippus gavi |
| status |
sp. nov. |
Hermippus gavi View in CoL sp. nov.
Type material. Holotype: Female ( ADSH 10901303): India, Kerala, Pathanamthitta, Gavi (9o26'09.07''N and 77o09'56.78''E), 3942 ft. alt., Pradeep M. S. leg. 21 December 2013, by hand.
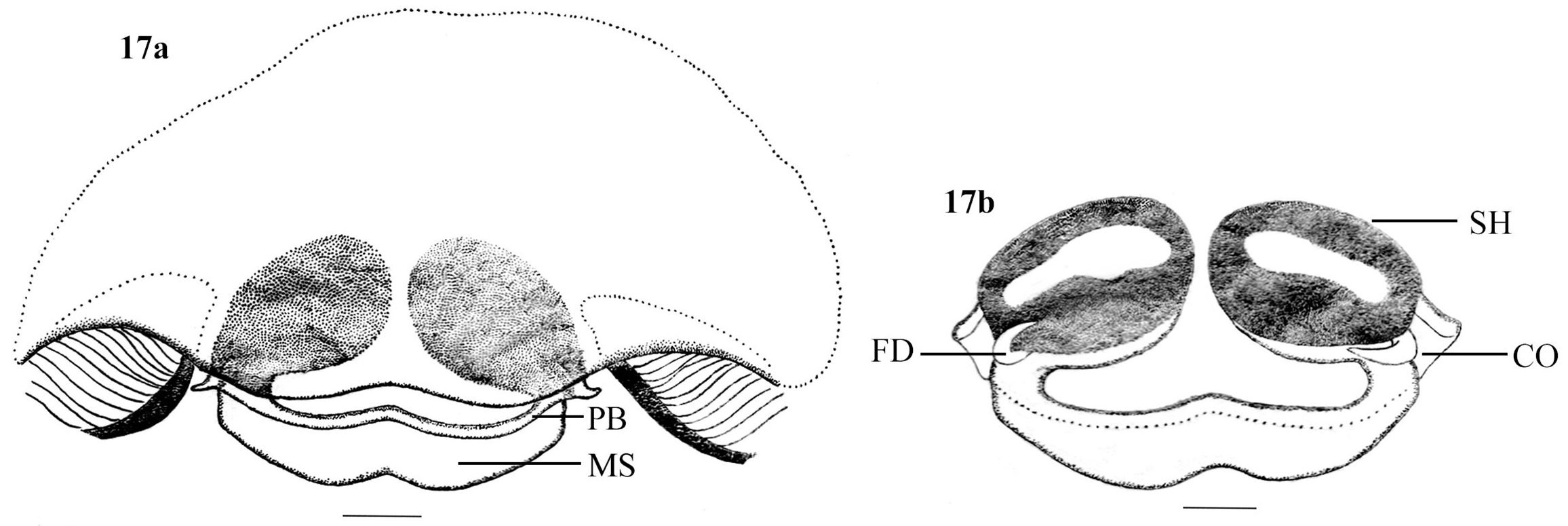
Diagnosis. Females of H. gavi sp. nov. can be recognized by the presence of highly sclerotized epigyne with ‘W’ shaped median septum (Figs 15a & 17a), the oval, obliquely placed spermathecae ( Fig 17 View FIGURES 17 a – 17 b b) and the characteristic dorsal opisthosomal pattern ( Figs 14a View FIGURES 14 a – 14 d, 15 a – 15 b & 16 & 20).
Description. FEMALE ( Figs 14a–14 View FIGURES 14 a – 14 d, 15 a – 15 b & 16 d, 16 & 20): Carapace uniform coquelicot, shiny, finely granulated, almost glabrous. Fovea longitudinal, reddish. Eyefield black. Clypeus black, finely granulated. Chilum triangular, wider than long, dark brown. Chelicerae black, promargin with two teeth, while retromargin without teeth or denticles. Labium and maxillae light brown; maxillae scopulated. Sternum yellowish-orange, posterior tip short, sharp, with poorly defined inter-coxal extensions. Opisthosoma oval, hairy, dorso-laterally black; dorsum provided with a characteristic pattern of chalk-white patches ( Figs 14a View FIGURES 14 a – 14 d, 15 a – 15 b & 16 & 20); lateral opisthosoma with a narrow, longitudinal, roughly zig-zag chalk-white stripe; venter grayish with a few chalk-white stripes. Spinnerets yellowish; spigots big and conspicuous (Fig 20). Posterior ventral spines present, arranged in two rows (Fig 16). Legs yellowish-orange; tarsi rebordered, with 4–7 short spines. Palpal segments yellowish-orange with dark brown tarsus. Body length 7.21. Prosoma length 3.48, prosoma width (at the middle) 2.73, prosoma thickness (at the middle) 2.04. Opisthosoma length 3.73, opisthosoma width (at the middle) 2.82, opisthosoma thickness (at the middle) 2.62. Eyes diameter: AME 0.16. ALE 0.22. PME 0.15. PLE 0.22. Eye interdistance: AME–AME 0.10.
PME–PME 0.25. AME–ALE 0.37. PME–PLE 0.50. AME–PME 0.16. ALE–PLE 0.08. Clypeus height at AMEs 0.76, at ALEs 0.56. Chilum length 0.17, width 0.42. Chelicera length 1.11. Measurements of palp and legs. Palp 2.54 [0.78, 0.51, 0.46, 0.79], I 8.72 [2.36, 1.01, 1.91, 2.02, 1.42], II 8.79 [2.43, 1.04, 1.85, 2.12, 1.35], III 8.47 [2.34, 1.02, 1.71, 2.24, 1.16], IV 10.04 [2.49, 1.05, 2.01, 2.96, 1.53]. Leg formula: 4213. Spination. Palp. femur 0 200, patella 1010, tibia 3213, tarsus 4054; legs: femora I–II 1300, III–IV 0500; patella I 0 0 0 0, II–IV 1000; Tibia I 1006, II 2016, III–IV 2136; metatarsus I 1016, II 1017, III 4044, IV 4048; Tarsi I–II 0 0 0 4, III 0 0 0 7, IV 0 0 0 6. Copulatory organ (Figs 15a, 15b, 17a & 17b): Epigyne simple and nearly rectangular (Figs 15a & 17a). Posterior border line of the epigyne thick with a median concavity (Figs 15a & 17a). Median septum W–shaped, highly sclerotized (Figs 15a & 17a). Spermathecae oval, obliquely placed, without definite copulatory duct or head (Figs 15b & 17b). Fertilization duct with a sharp proximal bent, lies in close contact with the proximo-lateral part of spermathecae ( Fig 17 View FIGURES 17 a – 17 b b).
Male. Unknown
Etymology. The specific epithet is a noun in apposition and is referring to the type locality of the species. Gender masculine.
Natural history. H. gavi sp. nov. inhabits semi-deciduous forest.
Distribution. At present known only from the type locality in Kerala, Southern India.
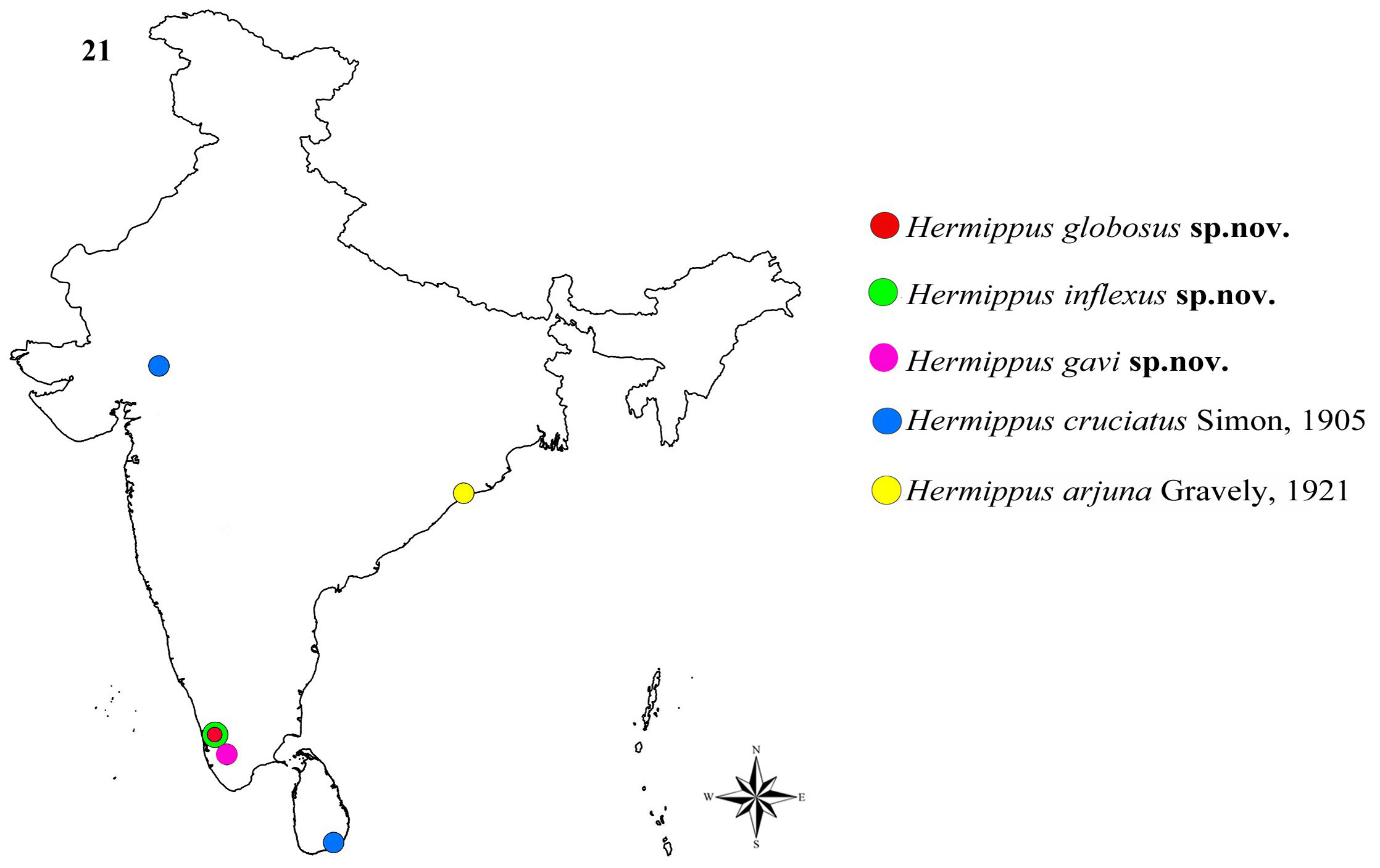
Note. All the described Oriental Hermippus species ( Fig 21 View FIGURE 21 ) can be separated from those of the Afrotropical species by four homoplasious characters: presence of posterior ventral spines (PVS) in front of the spinnerets, absence of external tooth on the cheliceral fang, absence of dorsal abdominal scutum and rebordered tarsi of legs. By considering these, all the described species of Hermippus can be categorized into two species groups:
The cruciatus View in CoL –group: Represented by all the five described Oriental species including H. cruciatus Simon, 1905 View in CoL , H. arjuna Gravely, 1921 View in CoL , H. globosus View in CoL sp. nov., H. inflexus View in CoL sp. nov., H. gavi View in CoL sp. nov.
The loricatus View in CoL –group: Represented by all the seven described Afrotropical species including H. loricatus Simon, 1893 View in CoL , H. affinis Strand, 1906 View in CoL , H. schoutedeni Lessert, 1938 View in CoL , H. septemguttatus Lawrence, 1942 View in CoL , H. minutus Jocqué, 1986 View in CoL , H. tenebrosus Jocqué, 1986 View in CoL and H. arcus Jocqué, 1989 View in CoL .
Two hypotheses can be suggested to explain the migration of the genus Hermippus View in CoL from Africa to India: one through Madagascar before its separation from India or through ‘Oman-Kohistan-Dras Island Arc’. If the first hypothesis is true, then Madagascar should have representatives of an African genus like Hermippus View in CoL . But Hermippus View in CoL species are not recorded so far from Madagascar. It is already hypothesized that migration of zodariid genera like Cydrela Thorell, 1873 View in CoL and Hermippus View in CoL from Africa to the Oriental Region might have occurred after the separation of Indian plate from Madagascar ( Dankittipakul et al. 2012). It is therefore plausible that the migration of Hermippus View in CoL spp. from Africa to Indian subcontinent occurred after its separation from Madagascar through the ‘Oman-Kohistan-Dras Island Arc’.
After its African origin, probably during the Early Cretaceous around 105 MYA after the separation of South America from Africa, Hermippus View in CoL species might have started their migration to Indian subcontinent around 65–60 MYA by the end of the Cretaceous during the Cretaceous–Tertiary (K/T) period. During this time, the Indian subcontinent, during its northward drift to Asia, reattached to northeast Africa through the land bridge formed of a volcanic island arc named Oman-Kohistan-Dras Island Arc, which may have provided the crucial biotic link between Africa and India during the Late Cretaceous ( Chatterjee & Scotese 2010, fig 7.3). The ‘Oman-Kohistan- Dras Island Arc’ hypothesis was proposed as an alternative for the ‘Greater Somalia’ hypothesis proposed by Briggs (2003), Chatterjee & Scotese (1999) and Chatterjee & Rudra (1996). The ‘Greater Somalia’ hypothesis was proposed to explain the existence of a continental corridor promoting faunal migration between India and Africa during the Late Cretaceous time. However, this hypothesis was rejected ( Chatterjee & Scotese 2010) as the ‘Greater Somalia’ broke up from Africa during the Late Triassic-Early Jurassic time due to the spreading of the Neotethys. Before the reattachment with Africa, Indian subcontinent had separated from Madagascar also. The cruciatus View in CoL —group of Hermippus View in CoL might have migrated from Africa to Indian subcontinent after its separation from Madagascar through the land bridge of Oman-Kohistan-Dras Island Arc. After reaching the Indian subcontinent, the cruciatus View in CoL —group of Hermippus View in CoL might have faced and survived the second and third phases of the fatal Deccan volcanic eruptions happened in the Indian subcontinent around 65 MYA, one of the major causative factors resulted in the Cretaceous–Paleogene (K/Pg) mass extinction ( Keller et al. 2009) and one of the species of the cruciatus View in CoL —group ( H. cruciatus Simon, 1905 View in CoL ) extended its habitat into Sri Lanka also ( Fig 21 View FIGURE 21 ).

| ADSH |
Arachnology Division, Sacred Heart College |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hermippus gavi
| Sankaran, Pradeep M., Jobi, Malamel J., Joseph, Mathew M. & Sebastian, Pothalil A. 2014 |
H. arcus Jocqué, 1989
| Jocque 1989 |
H. minutus Jocqué, 1986
| Jocque 1986 |
H. tenebrosus Jocqué, 1986
| Jocque 1986 |
H. septemguttatus
| Lawrence 1942 |
H. schoutedeni
| Lessert 1938 |
H. arjuna
| Gravely 1921 |
H. affinis
| Strand 1906 |
H. cruciatus
| Simon 1905 |
H. cruciatus
| Simon 1905 |
H. loricatus
| Simon 1893 |
Cydrela
| Thorell 1873 |