
Gonolobus caamali Carnevali & R. Duno, 2022
|
publication ID |
https://doi.org/10.11646/phytotaxa.530.2.2 |
|
DOI |
https://doi.org/10.5281/zenodo.5834549 |
|
persistent identifier |
https://treatment.plazi.org/id/03DEFD71-E42F-2658-AE9A-A420FDA9FECB |
|
treatment provided by |
Plazi |
|
scientific name |
Gonolobus caamali Carnevali & R. Duno |
| status |
sp. nov. |
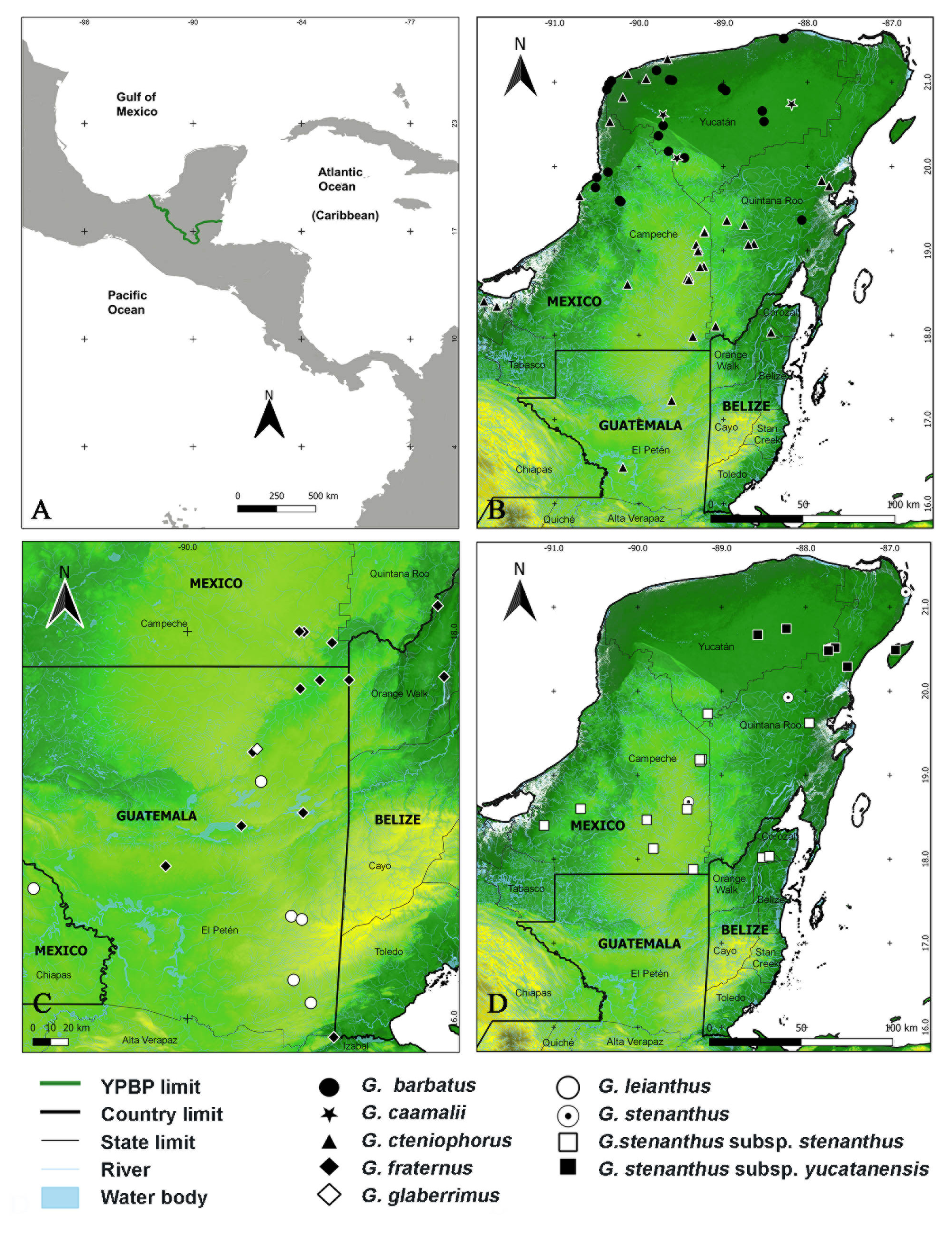
Gonolobus caamali Carnevali & R. Duno , sp. nov. ( Figs. 1B View FIGURE 1 , 4 View FIGURE 4 , 5 View FIGURE 5 ).
A species of Gonolobus phenetically similar and probably related to G. erianthus Decaisne (1844: 592) , a taxon that grows at higher elevations of (200–)900–1300(–1600) m in central Mexico south to Nicaragua in cloud forests, whereas the new species grows in lowland, seasonally dry forests at 10– 100 m. Furthermore, G. caamalii has oblong to oblong elliptic, narrower (3.0–3.2 times longer than wide) corolla lobes whereas they are proportionally wider (1.7–1.9 times longer than wide) in G. erianthus . Live flowers have strikingly different color patterns as G. caamalii has dark yellow green corolla lobes with a dark purple (almost black) longitudinal, callose band along the proximal 2/3 on the right side of the lobes whereas this band is absent or pale yellow, slightly evident in G. erianthus . The new species exhibits a 1.1–1.3 mm broad, annular, continuous dark purple-black band, the faucal annulus, surrounding the gynostemium and corona at the base of the corolla lobes. This band is narrower and either absent or pale green in G. erianthus .
Type:— MEXICO. Yucatán: Mun. Valladolid, 680–700 m al E del poblado de Chan Yokdzonot 2, unos 1380–1400 m al este del desvío Valladolid-Chan Yokdzonot , unos 3.7–4 km al norte en línea recta de Valladolid, 20°44’15.34”N, 88°11’23.46”W, 25 m, 24 November 2020; selva mediana subcaducifolia muy húmeda en esta época del año, G. Carnevali, R. Duno & M.A. Caamal Dzul 8573 (Holotype, CICY; isotypes, GH, MEXU, MO) GoogleMaps .
Herbaceous perennial vines, growth more of less monopodial, rarely branching. Latex white. Stems pubescent,with short and long, pale brown yellow, retrorsely spreading hairs up to 0.5 mm long, glandular capitate trichomes absent, pubescence dense throughout, not arranged in lines, denser distally and at the internodes, nodes densely pubescent, internodal colleters 3, 0.1 mm long. Leaf blades ovate, elliptic-ovate to oblong-ovate, 5.5–8.5(–11.6) × (3.0–)3.5– 6.0(–7.3) cm, apex acute to abruptly acuminate, acumen narrowly obtuse, 1.5–2.1 cm, bases always deeply cordate, lobes (0.7–)1.0–1.2(–1.5) cm long, convergent or more rarely somewhat parallel, often overlapping, sinus 1.0– 1.5 cm, adaxial surface very sparsely strigose, glandular capitate trichomes absent, eglandular trichomes scattered along the major and minor veins, margins entire with glandular capitate, hairs, 0.3 mm long, colleters 5, borne on each side of the petiole base, two in the base, two distal, and one in the apical, the basal one 0.5 mm long, the smaller one 0.1 mm long, eventually deciduous, abaxial surface paler than the adaxial surface, laxly pubescent throughout, nerving of 5–6 pairs of primary, the proximal retrorse and innerving the basal lobe, the rest at a 45° degree to the midnerve, not reaching the margin, nerves raised with a line of hairs; petioles (3.5–)4.0– 5.5 cm long, densely strigose with very short hairs throughout, more dense distally, becoming glabrescent proximally, glandular capitate trichomes absent; stipular colleters 2(–4), ca. 0.3 mm long, 1-2 borne on each side of the petiole base. Inflorescence subcapituliform, 1–3(–5)-flowered, the distal flowers maturing first; peduncles (1.5–)1.7–2.0 cm long, indument as on the stems but hairs shorter; rachis very short, 0.3–0.5 cm; pedicels 2.5–2.7(–3) cm long, indument as in the peduncle; floral bracts ca. 3 × 0.4 mm, eventually deciduous, pubescent as in the peduncles on both surfaces. Flowers showy, flat, presented vertically. Calyx lobes 5, 10–11 × 3–4 mm, ovate- elliptic to triangular elliptic, abruptly acuminate, margins pubescent, both surfaces strigose throughout, margin with longer hairs, glandular capitate trichomes absent, colleters 1-2 per sinus. Corolla 5-lobed, lobes green or yellowish green, with slightly darker, inconspicuous, reticulate nerving, and a dark-purple to almost black longitudinal, callose band along the proximal 2/3 on the right side of the lobes, the apex of the corolla lobes tinged in the same hue, the base of the corolla, surrounding the corona and the gynostemium, provided with a 1.1–1.3 mm broad, annular, continuous dark purple-black band of irregular width but wider opposite the anthers, strigose proximally with 0.02 mm long hairs; corolla lobes oblong-triangular to oblong, acute to narrowly acute, lobes 15–18 × 4.7–6 mm, wider proximally (5.8–6.0 mm wide), dextrogyrous at the distal half forming an angle of 30–35° to the general axis of the flower, lobes slightly overlapping proximally, glandular swelling absent in the sinus; adaxial surface glabrescent but densely hairy on the dark band, the pubescence mostly of eglandular trichomes to 0.4–0.9 mm long, abaxial surface moderately pubescent with eglandular trichomes 1.0 mm long, but conspicuously more pubescent near the apex, glabrescent on the underside of the callose, dark-colored band. Faucal annulus a distinctly raised ridge opposite each corolla lobe sinus, inconspicuously 5-lobed, 0.5–0.6 mm tall, with upper margins strigose with 1.2 mm long hairs. Gynostegial corona of fused staminal (Cs) and interstaminal parts, single, 5-lobed, 5 mm diameter, adnate to the ring, corona lobes 2 × 1.2 mm, pale green yellow, subquadrate reniform, somewhat bowl shaped, fleshy, smooth; gynostegium with stipe 2.5 mm high × 1.2 mm diameter, 5-winged, each wing with a small, fleshy projection distally; dorsal anther appendages 1.6–1.8 × 2.2–2.3 mm, deflexed, reniform-subquadrangular, wider proximally, shallowly bilobed at apex, thinly fleshy, concave, shiny wine-red; style head 3.2–3.8 mm wide, terminal appendages appressed to margin of style apex, guide rails +/- indistinct, 0.2–0.3 mm. Pollinarium with an ovoid corpusculum, 0.35–0.36 x 0.05–0.36 mm, dark brown, caudicle 0.5–0.6 × 0.3.0- 0.31 mm, flat, pollinia 1.6–1.7 x 0.6–0.7 mm, ovoid-ellipsoid apically rounded, translucent proximally, somewhat excavated ellipsoid, complanate. Follicles known only from a photograph, ovoid, sharply 5-winged, peduncles recurved.
Paratypes:— MEXICO. Yucatán: Mun. Abalá: 15 km al S del poblado de Yaxcopoil, a lo largo de la carretera Mérida-Muna , 20°36’30”N, 89°42’50”W (correctly 20°36’35.93”N, 89°42’52.60”W), approx. 12 m, “selva baja caducifolia, aguada o cenote de unos 200 m de diámetro; trepadora herbácea, rara localmente; látex blanco; flores verdes”, 7 November 2001, G. Carnevali, J.L. Tapia & F. May & M. Gómez 6438 ( CICY) GoogleMaps ; same locality, cultivated at Jardín Botánico Regional Roger Orellana, at the Centro de Investigación Científica de Yucatán, flowering 15 July 2015, G. Carnevali 8579 ( CICY, spirits) GoogleMaps . Mun. Oxkcutzcab, Helen Moyers Biocultural Reserve at Rancho Kiuic ( Kaxil Kiuic ), approx. 20°5.79’N, 89° 33.06’W, [90–100 m], “on the trail to Santa Rita, near the first haltun. Herbaceous liana; corolla green, contorted; gynostegium deep maroon”, 20 October 2005, W.J. Hayden 4923 ( URV) GoogleMaps . Mun. Tinum: Rancho San Pedro , 20°47´N, 88°23’W, 22 m, 24 November 1980 “flor verde negra”, E. Ucán Ek 569 ( CICY, XAL) GoogleMaps . Mun. Valladolid , same locality as type, 28 August 2020, G. Carnevali, R. Duno & M.A. Caamal Dzul 8550 ( CICY, SEL) GoogleMaps .
IUCN Conservation assessment:—EN. Gonolobus caamalii meets criterion B1 a, b (i, ii, iii, iv) for a Vulnerable species (V) and B2 a, b (i, ii, iii. iv) for Endangered species (EN). It is known from only four localities all in the state of Yucatán; its extent of occurrence is of 5,324.757 km 2, and the area of occupancy is of 16.0 km 2. EN B2 ab (i, ii, iii. iv).
Apparently, Gonolobus caamalii is an extremely rare species, having been found only four times at four different localities. At the type locality in northern Yucatán, we were able to record about 20 individuals on a dirt road along a span of 3–4 kilometers. Only a few specimens were in flower at any given time during the two visits we made at the locality. This population is severely threatened because of the imminent urban expansion of Valladolid, a medium sized city that is about 4 km south, and which is growing fast. The population SW of Abalá is severely threatened because of the expansion of cattle pastures and it is expected to disappear in the near future. The same applies to the Tinum locality. The population at the Helen Moyers Biocultural Reserve at Rancho Kiuic is protected and it is at this locality where the best survival expectations for the species are met. Because the species is rare, both its EOO and AOO are small, only occurs naturally in nearly undisturbed vegetation, and most of its known populations occur in areas where the original vegetation is currently being or is likely to be substituted by anthropogenic ecosystems during the next few years, we propose the species to be treated as Endangered (EN).
Eponymy: Named after Miguel Ángel Caamal Dzul, of Chan Yokdzonot 2, Yucatán, who collected for the second time this interesting species. He has a keen eye for rare plants and enthusiastically collects, cultivates, and photographs the native flora.
Notes:— Gonolobus caamalii is similar and putatively related to G. erianthus , yet both taxa are amply different in several relevant aspects. Both species have green corollas with reticulate nerving of relatively similar shapes and with one of the margins crenulate, a yellow corona, and reddish anther appendages. However, our new taxon is a lowland species occurring at elevations of 10–100 m in tropical dry forests whereas G. erianthus grows at higher elevations of (200–)900–1300(–1600) m in central Mexico south to Nicaragua in tropical humid to cloud forests. The corollas of both species, albeit relatively similar, are readily diagnosed. In G. caamalii , the corolla lobes are asymmetrically oblong to oblong elliptic, 15–18 × 4.7–6 mm and relatively narrow (3.0–3.2 times longer than wide). On the other hand, G. erianthus features asymmetrically elliptic corolla lobes 15–23 × 9–12 mm that are proportionally wider (1.67–1.9 times longer than wide). Moreover, the color patterns of live flowers (even somewhat noticeable in pressed material) are unmistakable. Gonolobus caamalii has dark yellow green corolla lobes that feature a dark purple (almost black) longitudinal, callose band along the proximal 2/3 on the right side of the lobes; this band is absent, or it is yellow and only slightly evident in G. erianthus . In addition, there is 1.1–1.3 mm broad, annular, continuous dark purpleblack band surrounding the gynostemium and corona at the base of the corolla lobes. This band is narrower and either absent or dark green in G. erianthus . Gonolobus caamalii is fairly distinctive in possessing a very dark, almost black brownish purple, callose longitudinal band parallel to the right margin of the petals and in the fact that the petals are sharply tilted dextrogyrously forming an angle of 30–35° to the general axis of the flower. In Stevens (2010) useful key to the Mesoamerican Gonolobus , G. caamalii would key out to couplet 36b because of its abaxially moderately pubescent corolla of 15–18 mm long, its uniformly pubescent internodes, and the anther appendages lacking 2 distinct lobes or “arms”. Then, it would key out to G. erianthus in couplet 37a. However, G. caamalii is sharply distinct from G. erianthus (a species ranging widely from Nuevo León and Nayarit, Mexico, to Nicaragua) in several respects as discussed above. In the YPBP, it is difficult to confuse it with any other species.
The species is known only from a narrow, arch-like band of seasonally dry forests under Ax(Wo) climate according to the Köppen system (as modified by García, 2004 ). Here, rains occur mostly in the summer and precipitation is in the 1000–1200 mm range. The vegetation is locally called “selva mediana subcaducifolia” (“medium height semideciduous forest”)and is structurally dominated by several species of woody deciduous species such as Jatropha gaumeri Greenman (1907: 256) , Margaritaria nobilis Linnaeus (1781: 428) (Euphorbiaceae) , Bursera simaruba ( Linnaeus 1753a: 1026) Sargent (1890: 260) (Burseraceae) , Leucaena leucocephala ( Lamarck 1783: 12) de Wit (1961: 54) subsp. leucocephala , Caesalpinia gaumeri Greenman (1912: 330) , Bauhinia ungulata Linnaeus (1753b: 374) , Gliricidia maculata ( Kunth 1823: 393) Kunth ex Walpers (1842: 679) (Fabaceae) , Cedrela odorata Linnaeus (1759: 940) (Meliaceae) , Ceiba aesculifolia ( Kunth 1821: 298) Britten & Baker (1896: 175) , Hampea trilobata Standley (1923: 787) (Malvaceae) , and Gymnopodium floribundum Rolfe (1901 : t. 2699) ( Polygonaceae ). The area is particularly rich in scandent members of the Apocynaceae with species such as Matelea stenosepala Lundell (1942: 398) , Dictyanthus yucatanensis Standley (1930: 37) , Cynanchum rensonii Woodson (1941a: 210) , Macroscepis diademata (Ker Gawl. in Edwards 1817: 3, t. 252) Stevens (1983: 405), Echites yucatanensis Millspaugh ex Standley (1930: 35) , and Mandevilla subsagittata ( Ruiz & Pavón 1799: 19) Woodson (1932: 60-70) . Herbs on the forest floor include Agave petskinil (Orellana et al., in Hernández et al. 2008: 173) Thiede (2012: 94) ( Asparagaceae ), Oeceoclades maculata ( Lindley 1821: pl. 15) Lindley (1833: 237), Sarcoglottis sceptrodes (Reichenbach f. 1855: 214) Schlechter (1920: 421) ( Orchidaceae ), and Hybanthus thiemei ( Donnell Smith 1905: 1) Morton (1944: 81) (Violaceae) .
The northern portion of the YPBP seems to be a center of endemism for the Gonolobinae with at least 10 species restricted to this area, including Gonolobus caamalii and the recently described Matelea falcata Juárez-Jaimes et al. (2021: 113) . It seems likely that several other novelties are yet to be discovered in this area. Most of these endemic Gonolobinae consist of small populations, most likely threatened, surviving in remnants of the once extensive semideciduous forests that covered the Yucatan Peninsula before pervasive “slash and burn” agriculture removed most of them.
Gonolobus stenanthus subsp. yucatanensis has been collected near and at the type locality of G. caamalii . For example, G. Carnevali et al. 8377 (CICY, CHIP, CIQR, F, FCME, G, GH, MEXU, MO, SEL, UADY, UCAM, UJAT, XAL; Fig. 3I View FIGURE 3 ) was collected less than 4.5 km to the NW whereas C.L. Lundell & A.A. Lundell 7577 (GH, MEXU, MICH) was collected 41 km E of the type collection of G. caamalii . More recently, M.A. Caamal 01 (CICY; Fig. 3H View FIGURE 3 ) was collected a few hundred meters for the type locality. Thus, it is evident that both species are sympatric or parapatric in the area. However, there is no morphological evidence of gene flow between the two very distinct Gonolobus species. Aside from possible morphological or genetic barriers to gene flow between both taxa, G. stenanthus subsp. yucatanensis flowers at the end of the dry season or beginning of the rainy season (April-July) whereas G. caamalii , at least under natural conditions, blooms toward the end of the rainy season (October-November) providing an additional prezygotic barrier to gene exchange.



| CICY |
Centro de Investigación Científica de Yucatán, A.C. (CICY) |
| URV |
University of Richmond |
| XAL |
Instituto de Ecología, A.C. |
| SEL |
Marie Selby Botanical Gardens |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Asclepiadoideae |
|
Genus |