
Kamopanorpa rotunda, BASHKUEV & SUKATSHEVA, 2021
|
publication ID |
https://doi.org/ 10.11646/palaeoentomology.4.5.11 |
|
publication LSID |
lsid:zoobank.org:pub:50F1EB3E-450C-4B2C-A4F8-11E113C0FD02 |
|
persistent identifier |
https://treatment.plazi.org/id/03D8B42B-F236-FF9B-FF65-F926CC06F7D0 |
|
treatment provided by |
Plazi |
|
scientific name |
Kamopanorpa rotunda |
| status |
sp. nov. |
Kamopanorpa rotunda sp. nov.
( Figs 1C View FIGURE 1 , 2C View FIGURE 2 )
Type material. Holotype PIN 5386 View Materials /5±, well-preserved forewing with anal area missing.
Etymology. From Latin rotundus (round), referring to roundish shape of wing.
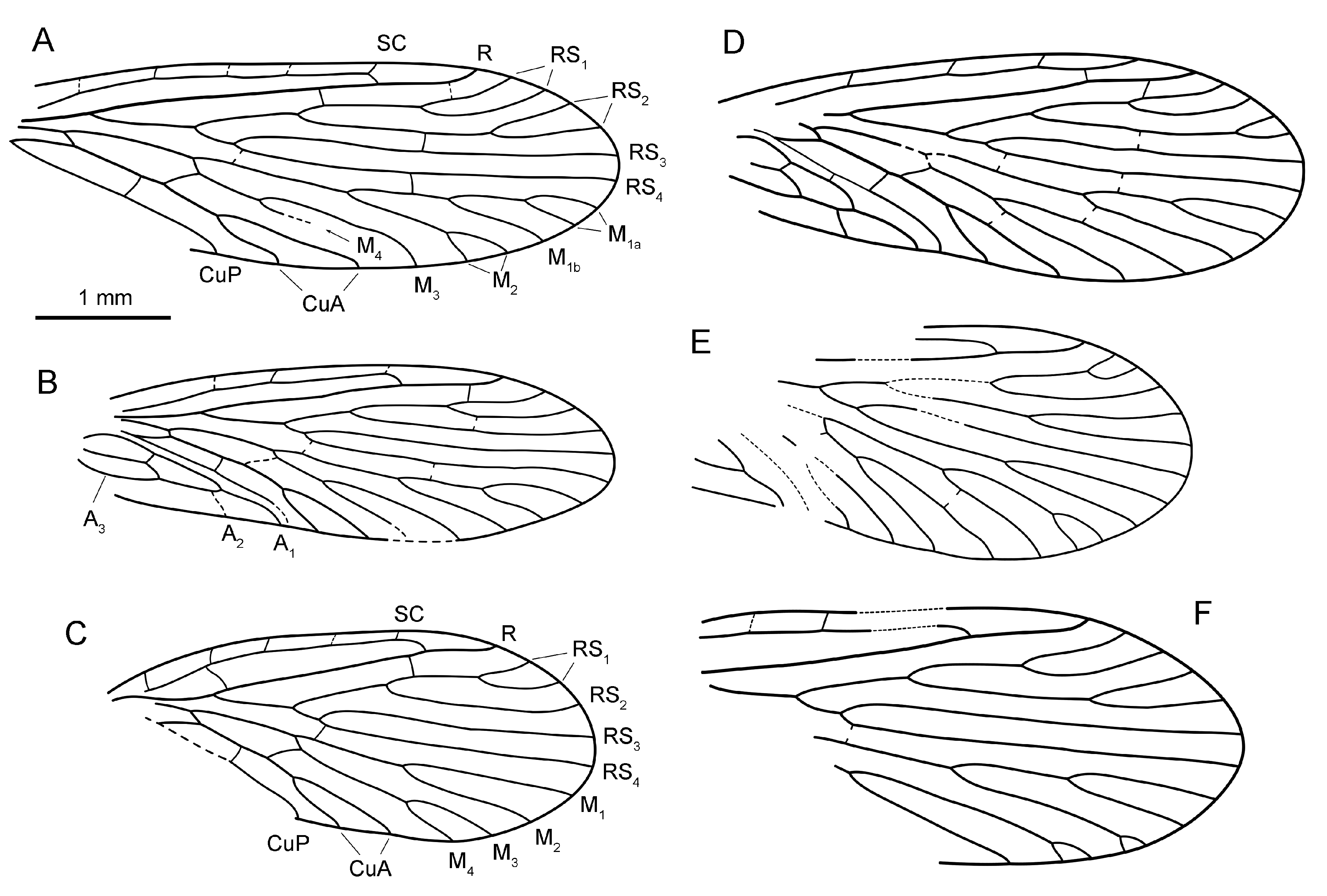
Diagnosis. Wing short and rounded; SC ending at R before level of RS 1 fork, with four subcostal crossveins
(besides apical one); subcostal space broad with long sigmoid crossvein. Clearly differs from all congeners by simplified venation of M with only four branches and very short M 3+4 fork.
Locality and horizon. Izykh Coal Mines, quarry no. 1, south of Belyi Yar , Khakassia, Minusinsk Coal
Basin, South Siberia; Izykh Formation, Middle Permian.
Description. Wing short, broadly ovoid (L:W ratio
1:2.2) with rounded apex at RS 3 –RS 4; anterior margin convex at 1/3 of wing length. SC reaching 2/3 of wing length, visually ending at R with short and weak veinlet to costal margin. Besides that apical vein, there are four short subcostal crossveins at equidistance from one another. Subcostal space broad, crossed by strong sigmoid crossvein sc–r at level of R–RS forking point.
Pterostigma not preserved. R curved basally, without pterostigmal branches. RS forking well before middle of wing, with five branches. Cells DC and MC open. M with four branches; M 3+4 forking very late, distal to M 1+2 fork. Base of CuA long, strongly oblique, M 5 much shorter. CuP preserved only by its distal half after cua-cup crossvein,
anal area missing. Crossveins sc–r, r–rs 1, m 3+4 –cua, and cua–cup distinct, others completely invisible, except for very faint rs–m.
Measurements. Forewing length, 3.6 mm, width,
1.6 mm.
Other material examined
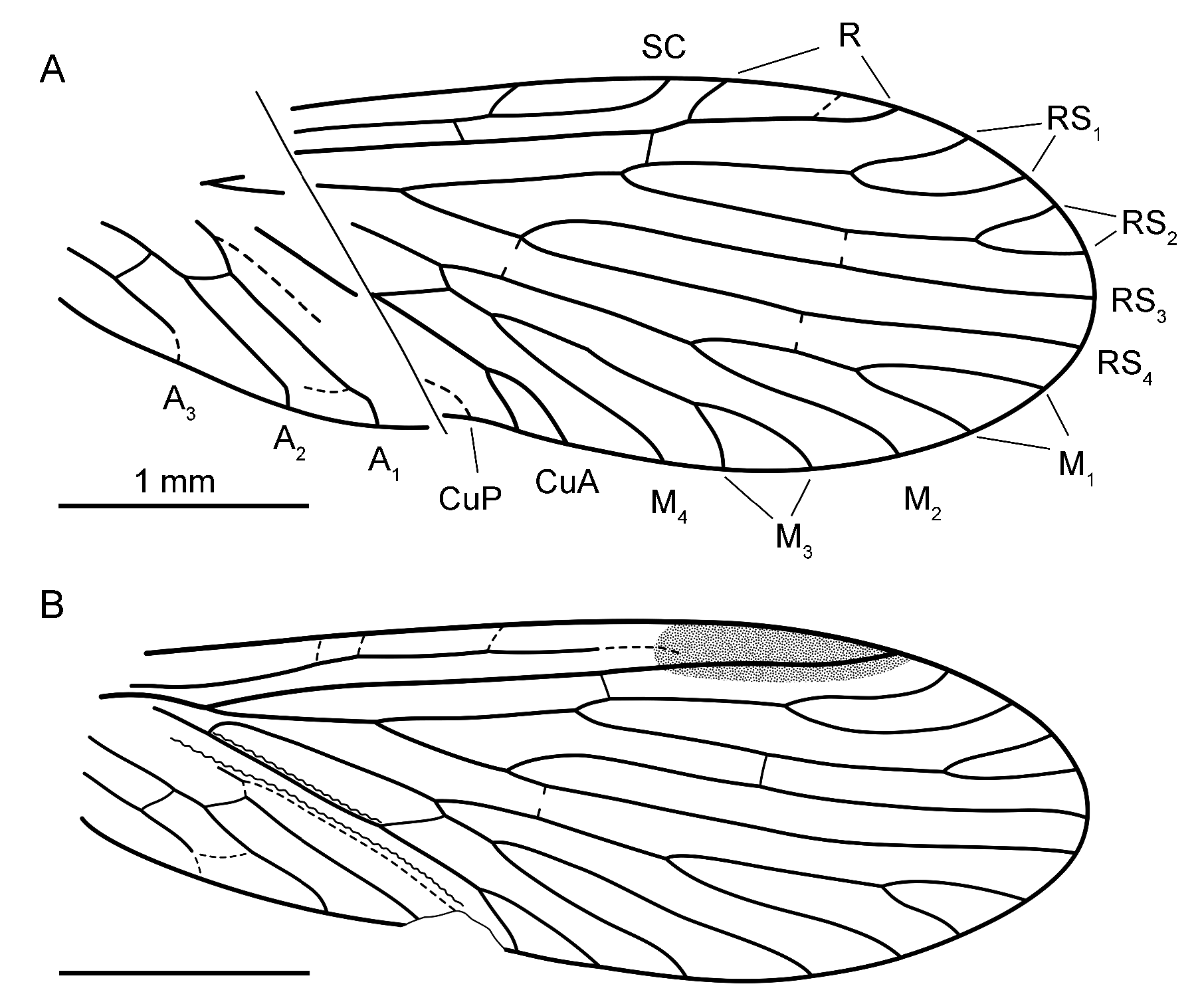
In addition to the described species, there are two wellpreserved hindwings, PIN 5386/2± and 10± ( Fig. 3 View FIGURE 3 ), and also several forewings (PIN 5386/4, 6±, 7±, 8±, 11±, 12, 13±) that are too fragmentary or poorly preserved to be referred to any particular species. Some of them show additional forkings on RS and M branches, notably
2
5386/8± ( Fig. 1E View FIGURE 1 ), which has RS with 8 branches in total with deep fork on RS 4 (the latter is only rarely found in Kamopanorpa: Novokshonov, 1992, 1993b ).
In microptysmatid hindwings, as was shown by Novokshonov (1997: 94), the anal veins have a tendency to looping, changing from freely reaching wing margin in Early Permian forms to coalesced apically and forming distinct anal loop in the Late Permian ones.Re-examination of the hindwing of K. pritykinae ( Sukatsheva, 1976) has shown that A 2+3 does not just end blindly in membrane but its unsclerotized apical portion terminates on A 1 ( Fig. 4D View FIGURE 4 ). The new specimens show an arrangement of anal veins of transitional nature: the A 2 and A 3 meet inner wing margin and each is distally connected to the next anal by a very weak curved crossvein. Apical part of A 3 is unsclerotized in both specimens ( Figs 3 View FIGURE 3 , 4B, E, F View FIGURE 4 ).
An interesting detail was observed in specimen 5386/10±. On the posterior side of subcosta along its basal part up to the first crossvein, there is a dense row of sockets (bases of setae), directed basad and upwards, which in all probability was a part of a wing coupling apparatus (the first direct evidence for wing coupling in Palaeozoic amphiesmenopterans) ( Fig. 4C View FIGURE 4 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.