
Pliciloricus apteryx, Sørensen & Grzelak & Kristensen & Herranz, 2022
|
publication ID |
https://doi.org/ 10.1016/j.jcz.2022.06.004 |
|
publication LSID |
lsid:zoobank.org:pub:D67E1882-834D-469A-B034-B31D4C799533 |
|
persistent identifier |
https://treatment.plazi.org/id/7FDA2372-F12B-4D61-AD75-98BB83472D7D |
|
taxon LSID |
lsid:zoobank.org:act:7FDA2372-F12B-4D61-AD75-98BB83472D7D |
|
treatment provided by |
Felipe |
|
scientific name |
Pliciloricus apteryx |
| status |
sp. nov. |
3.1. Pliciloricus apteryx View in CoL sp. nov
( Figs. 2–5 View Fig View Fig View Fig View Fig ; Table 1).
urn:lsid:zoobank.org:act:7FDA2372-F12B-4D61-AD75-98BB83472D7D
Taxonomic account.
Phylum Loricifera Kristensen, 1983 View in CoL .
Family Pliciloricidae Higgins & Kristensen, 1986 View in CoL .
Genus Pliciloricus Higgins & Kristensen, 1986 View in CoL .
3.1.1. Pliciloricus apteryx View in CoL sp. nov. diagnosis
Adult Pliciloricus with broad mouth cone, extending into mouth tube with buccal tube forming projecting tongue-like structure. Introvert Row 1: six regular, club-shaped clavoscalids with 25 annulated rings in distal 1/3; two ventral clavoscalids strongly modified into thin and flexible scalids with only four annulated rings, about 1/3 from distal tip. Introvert Row 2: three regular spinoscalids alternating with four leg-shaped scalids with swollen bases and three “finger joint-like” constriction 1/3 from tip; distal 1/3 of scalids curved; double organ fused at proximal 1/3 only, extending into long, tapering distal tips that bend in a 45 ◦ angle and cross each other. Introvert Row 4: fifteen claw-like scalids with serrated shafts, and hawkbill blade-shaped tips. Neck with eight single trichoscalids posterior to two basal plates, and seven double trichoscalids, arranged side-by-side, posterior to three basal plates. Lorica with 22 plicae inclusive broad midventral plica. Crescentic anus forms the anterior margin of distinct, cuticular posteroventral rosette. Males with paired testes, connecting with terminal gonopore; female morphology unknown.
3.1.2. Etymology
Since this species is the first loriciferan described from New Zealand, we nick-named it “the kiwi loriciferan”. The species name, ‘apteryx ’, refers to the genus name Apteryx – the flightless kiwi bird, endemic to New Zealand.
3.1.3. Examined material
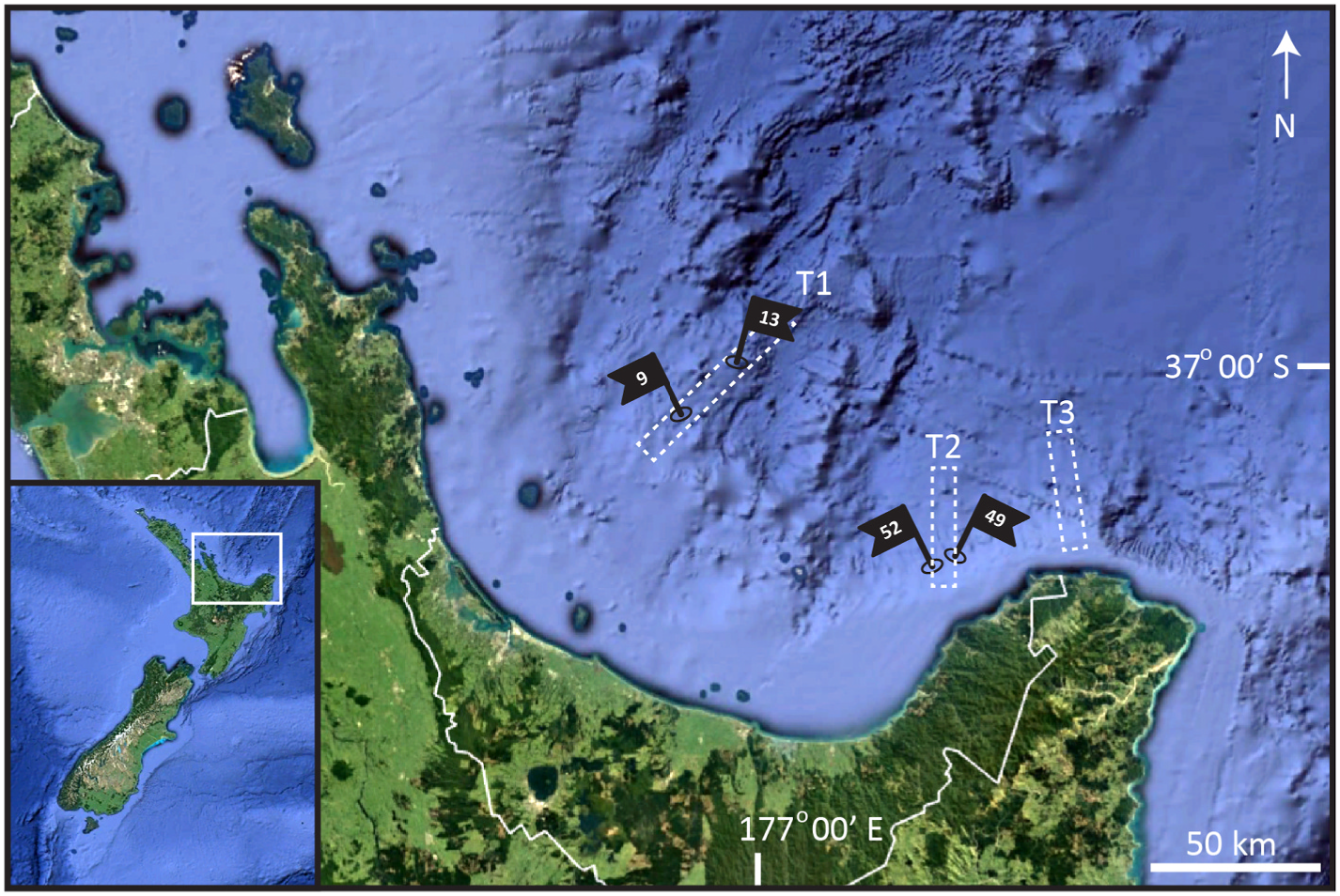
Holotype, adult male, collected from silt in April 2012, at St. 49 (sensu Rosli et al. 2016) at 1004 m depth on the continental slope, in the eastern part of Bay of Plenty, North Island , New Zealand (37 ◦ 29.90’N, 177 ◦ 37.55’E), mounted in glycerine, and deposited at NIWA under catalogue number NIWA-159431 GoogleMaps . Paratypes include one adult male (NIWA-159432), collected at same station as holotype, and three Higgins larvae collected at St. 9 and 13 (see Table 1 for further details). The male paratype is mounted at the same slide as the holotype, and the two Higgins larvae from St. 13 are also mounted together. Paratypes are deposited at NHMD under catalogue numbers NHMD-916683 to 916685. See Fig. 1 View Fig for localities and Table 1 for detailed station data .
3.1.4. Description of adult male
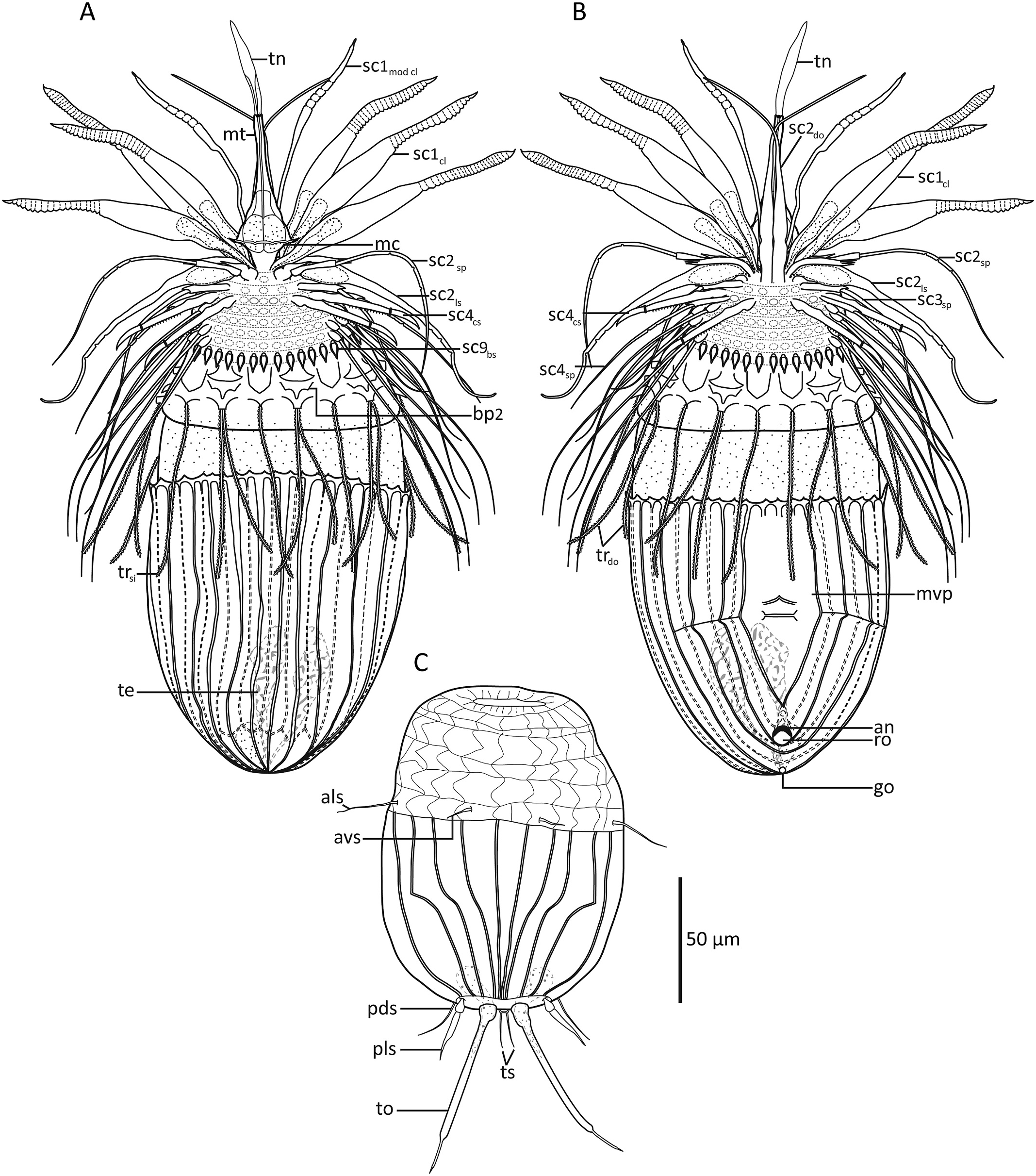
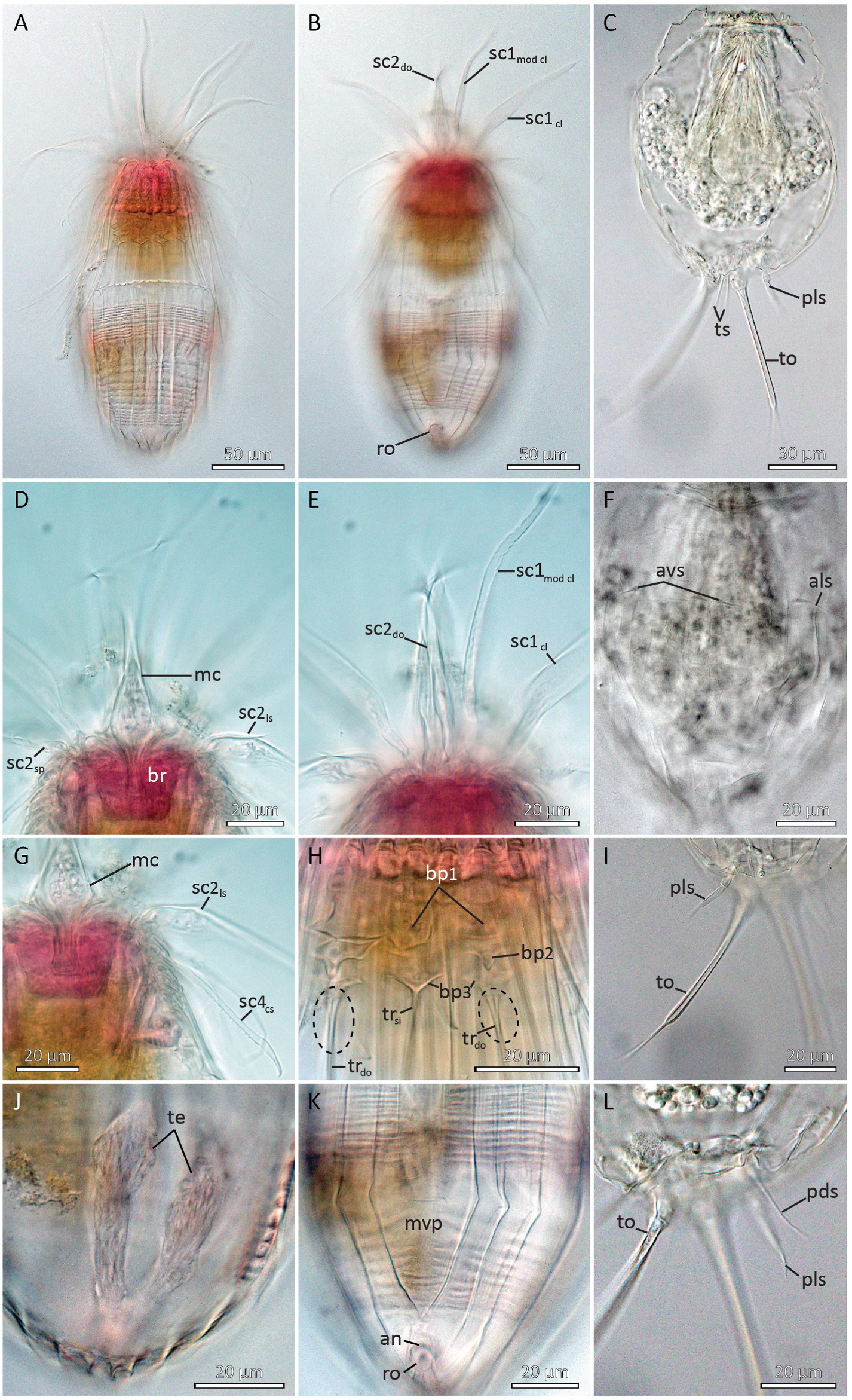
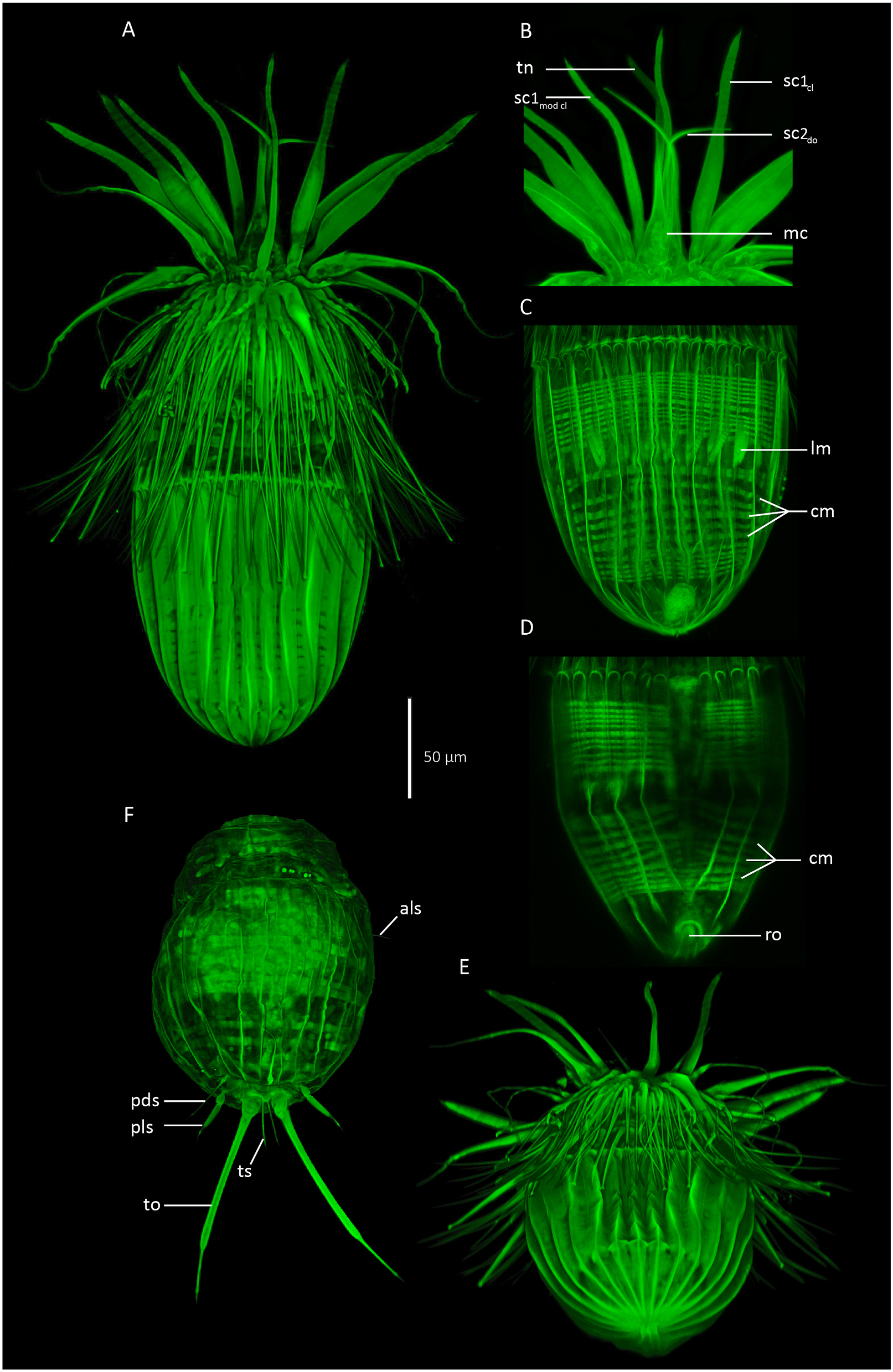
Holotypic male ( Fig. 2A and B View Fig , 3 View Fig , 4 View Fig A-B, D-E, G-H, J-K, 5A, C-E) measures 267 μm in length, including mouth cone, and 103 μm in width at its widest point, i.e., near the anterior lorica margin. The mouth cone measures 55 μm in length. It has a narrow base, but broadens conspicuously in the area around the pharynx, before narrowing again towards the mouth tube ( Figs. 2A View Fig , 4D View Fig and 5B View Fig ). The pharynx does not have any distinct internal structures, and no placoids or oral ridges were observed. A narrow buccal tube is visible inside the mouth tube, and a spatulate tongue-like structure projects from the mouth opening ( Fig. 5B View Fig ).
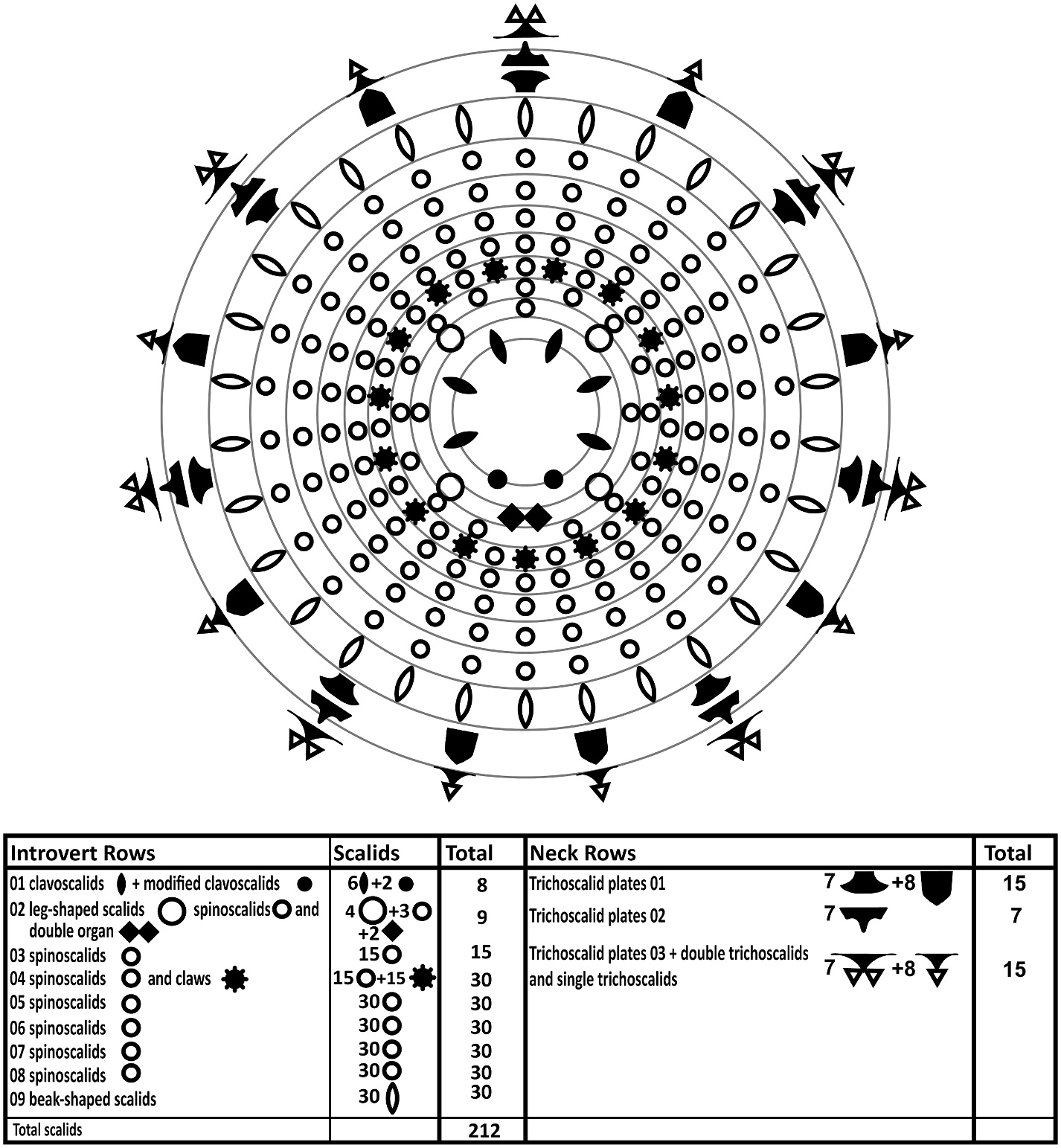
The introvert has nine rows of scalids ( Fig. 3 View Fig ). The anteriormost Row 1 consists of six regular, club-shaped clavoscalids and two much thinner, ventral modified clavoscalids. The regular clavoscalids are about 110 μm long, and consist of a narrow base, a smooth, flattened part representing 2/3 of the total scalid length, and a distal end-piece with about 25 annulations. The clavoscalids terminate into a small, anteriorly bent hook. The ventral modified clavoscalids are nearly as long as the regular clavoscalids, but rounded in cross-section. Each modified clavoscalid is also composed of three parts: a thin and flexible proximal 2/3, a slightly thicker part with four annulations, and a smooth end-piece that tapers abruptly at its tip ( Fig. 2A and B View Fig , 3 View Fig , 4B, E View Fig , 5B View Fig ).
Row 2 consists of four leg-shaped scalids, three more regular spinoscalids, and two scalids fused into a ventral double organ. Each leg-shaped scalid (length: 115 μm) consists of a swollen proximal part that terminates into a short fringe ( Fig. 4D View Fig ). The swollen basis narrows into a smooth mid-piece. About 1/3 from its distal tip, the leg-shaped scalid has three partial constrictions, resembling the joints in a human finger. The distal tip, ca. 1/4 of the scalid, is smooth and anteriorly curved. The three regular spinoscalids (length: 117 μm) alternate with the leg-shaped scalids and are located middorsally and midlaterally on the introvert. Each spinoscalid is composed of three units: a short, proximal, ball-shaped basis with an anterior spike, a short but thick, cylindrical mid-piece, and a long, acicular end-piece, representing 3/4 of the full scalid length. The end-piece has three, “finger joint-like” constrictions, and a pointed, non-curved tip. The double organ consists of two appendages (length: 88 μm) that are fused along their proximal 1/ 3. The fused parts of the appendages are relatively thick, but distally they narrow gradually into long, acicular tips that bend in a ca 45 ◦ angle and cross each other ( Fig. 2A and B View Fig , 3 View Fig , 4B View Fig , D-E, G, 5B).
Row 3 consists of fifteen uniform spinoscalids (length: 78 μm), composed of a short cylindrical basis and a long, smooth acicular end-piece ( Fig. 2A and B View Fig , 3 View Fig ).
Row 4 consists of fifteen regular spinoscalids alternating with fifteen claw-like scalids. The spinoscalids (length: 79 μm) are composed of a short cylindrical basis, and a long acicular end-piece with three “finger joint-like” constrictions in its proximal 1/3. Each claw-like scalid (length: 53 μm) is composed of a long shaft (3/4 of scalid length) with fine serration along its inferior margin, and a distal, hawkbill-shaped blade ( Fig. 2A and B View Fig , 3 View Fig , 4G View Fig ).
Rows 5 to 8 spinoscalids are very uniform. Each row has thirty spinoscalids (lengths: 80–110 μm), each consisting of a short ovate basis, and a long, smooth acicular end-piece. Minute plates alternating with spinoscalids in row 8, as found in certain congeners, not observed ( Fig. 2A and B View Fig , 3 View Fig ).
Row 9 consists of thirty, short beak-shaped scalids (length: 14 μm) ( Fig. 2A and B View Fig , 3 View Fig ).
The neck has eight single and seven double trichoscalids ( Fig. 3 View Fig ). The trichoscalids (lengths: 70–78 μm) are flattened with numerous small, triangular wings along their margins. The most ventral trichoscalids are a pair of singles, but otherwise the trichoscalids alternate between singles and doubles. Each single trichoscalid is located posterior to two basal plates. The anteriormost basal plate is elongate and pointed posteromedially. The posteriormost plate is chevron-shaped, with the single trichoscalid attaching at the tip of the chevron. Each double trichoscalid is located posterior to three basal plates. The anteriormost one is shaped like the traditional Inuit ulu-knife, with a broad blade on short medial stalk, whereas the median plate has two short lateral projections and a longer posteromedial projection. The posteriormost plate tends to be chevron-shaped, but has a small central basis instead of tip. The double trichoscalids attach side-by-side at this central basis ( Fig. 2A and B View Fig , 3 View Fig , 4H View Fig ).
The thorax (length ca 20 μm) has no appendages and apparently no plates, folds or other cuticular structures ( Fig. 2A and B View Fig , 4 View Fig A-B, 5A).
The abdomen is covered by a lorica with ca forty short, irregular spikes along its anterior margin. The lorica consists of 22 plicae, separated by primary double ridges, including the large midventral plica ( Fig. 2A and B View Fig , 4 View Fig A-B, K, 5A, C-D). Each plica consists of two, anteriorly arched ridges, separated by intracuticular secondary double ridges. All plicae on the dorsal side narrow and curve posteriorly, and meet each other in a terminal point close to the gonopore ( Fig. 5E View Fig ). The secondary double ridges of the five dorsal-most plicae split near the terminal end of the lorica, and expand in a postero-diagonal direction until they join the primary double ridges. This gives the posterior parts of these five plicae a diamond-shaped substructure ( Figs. 2A View Fig and 5A, E View Fig ). The lateroventral plicae also curve towards the terminal lorica tip, whereas the more ventral plicae curve even more and meet with their own counter-plicae on the posteroventral part of the lorica ( Figs. 2B View Fig and 4B, K View Fig , 5D View Fig ). The midventral plica is 30 μm broad at it widest point, and appears, based on the four anterior arches, to be formed by two fused plicae. Two transverse double ridges are present medially on the ventral plate: The anteriormost ridges form an anteriorly pointing chevron, whereas the posterior ones form a simple, straight fold. A transverse single ridge, horizontally aligned with this straight, midventral fold, expands through the three ventrolateral plicae next to the midventral one ( Figs. 2B View Fig and 4K View Fig ).
The anus is located ventrally, at the point where the two most ventrolateral plicae meet. The anal opening is crescentic and represents the anteriormost part of a cuticularised rosette structure. The gonopore is located posterior to the rosette, very close to the terminal point of the lorica ( Figs. 2B View Fig and 4B, K View Fig , 5D View Fig ). Flosculi were not observed in any part of the lorica.
Internally, attached to the cuticle, there are numerous circular muscles (ca 23 rings) encircling ten pairs of dorsal and eight pairs of ventral, longitudinal retractors. Elongate, paired testes are present in the posterior half of the lorica. The paired testis ducts meet at the gonopore, and they may or may not connect with a small, globular gland, located ventral to the testes, and very close to the gonopore ( Fig. 2A and B View Fig , 4J View Fig , 5 View Fig C-D).
3.1.5. Description of Higgins larva
All paratypic Higgins larvae were strongly contracted, and it is therefore impossible to provide any information about the head morphology. The trunk of the Higgins larva is divided into a thorax region and a loricated abdomen ( Figs. 2C View Fig , 4C View Fig and 5F View Fig ). The thorax has six transverse folds and numerous (> 22) longitudinal folds, giving it an irregular, chequer-patterned appearance. Two pairs of simple, acicular, unbranched setae are present between the two posteriormost folds in lateral and more ventral positions ( Figs. 2C View Fig and 4C, F View Fig , 5F View Fig ). Anterolateral setae measure 24 μm, whereas the anteroventral ones are much shorter, measuring only 11 μm.
The lorica has 22 plicae, separated by longitudinal folds. Dorsal folds are all strictly longitudinal in their orientation, whereas ventrolateral folds tend to slightly deviate in a more lateral direction, before they more anteriorly curve back and continue in a longitudinal arrangement ( Fig. 2C View Fig ). Three pairs of setae are located in the posterior part of the lorica ( Figs. 2C View Fig and 4C, I, L View Fig , 5F View Fig ). The posterodorsal setae (length: 24 μm), and the terminal setae (length: 15 μm) that attach between the toes, are all simple and acicular. The posterolateral setae (length: 26 μm) differ by having swollen bases, and a thick proximal part that tapers abruptly about 1/3 from the tip.
The toes are long and slender (total toe length: 78 μm), and divided into three portions: short swollen bases, a long, slender mid-piece (length: 55 μm), and abruptly narrowing tips (length: 16 μm) that terminate into even thinner, 2 μm long nibs ( Figs. 2C View Fig and 4C, I View Fig , 5F View Fig ). Internally, the proximal thirds of the toes are filled with glandular tissue that continue and expands into glands in the posterior part of the trunk. Openings of the toe glands were not observed, but since the glands do not extend into the offset tips, openings are expected to be present near the abrupt narrowing between the mid-piece and distal tips.
3.1.6. Notes on diagnostic features in adults
The combined morphology of the different head appendages is unique for P. apteryx sp. nov., but the species also shows certain character traits that are unique in themselves. These include the tongue-like structure protruding from the mouth tube, the clavoscalid and double organ morphology, and the arrangement of the double trichoscalids.
The tongue-like structure that extends from the mouth tube may either be an extension of the buccal tube or a separate, and perhaps retractable, structure inside the mouth tube. A structure like this has never been reported from any other loriciferan species, but it is currently impossible to determine if it is unique for Pliciloricus apteryx sp. nov. or simply has been overlooked in previous descriptions. Due to its extremely delicate nature, the tongue-like structure could not be visualised with conventional light microscopy or DIC, and we were unable to realise its presence until we examined the specimens with CLSM ( Fig. 5B View Fig ). Thus, we cannot reject that other species, which so far have been subject to LM examinations only, might have similar structures. However, we would expect the structures to be visible with SEM.
Species of Pliciloricus show much morphological variation in their clavoscalid morphology. The number of annulated rings in the clavoscalids varies between species, and so does the exact position of these rings. For instance, species like Pliciloricus pedicularis Gad, 2005 and P. senicirrus Gad, 2005 have numerous (+20) rings distributed throughout the entire length of each clavoscalid ( Gad 2005b; 2005c), whereas P. cavernicola and Pliciloricus hadalis Kristensen & Shirayama, 1988 have only four and three rings, respectively, concentrated in the distal third of each scalid ( Kristensen & Shirayama 1988; Heiner et al., 2009). The clavoscalids of P. apteryx sp. nov. have about 25 annulated rings, which is a relatively high number compared to most other species. However, opposed to all other species with more than ten rings, all annulated rings in P. apteryx sp. nov. are concentrated in the distal 1/3 of each scalid. Thus, we consider the clavoscalid morphology as diagnostic for the new species.
Also the double organ, located ventrally in introvert scalid Row 2 has a unique morphology. In most species of Pliciloricus the double organ consists of two, basally fused scalids. The fused parts of the double organ are slightly thicker than the free, distal ends, but the difference in thickness is usually not conspicuous. The unfused, distal tips tend to be straight and rigid (thus, pointing the same direction as the basal parts), equipped with setae or teeth, and terminating in an off-set tip or hook (see, e.g., P. enigmaticus Higgins & Kristensen, 1986 , P. leocaudatus Heiner & Kristensen, 2005 , and P. senicirrus ( Higgins & Kristensen 1986; Gad 2005c; Heiner & Kristensen 2005),). However, in P. apteryx sp. nov. the distal, unfused tips gradually extend into long, acicular appendages, and about halfway from their tips, they bend and cross each other ( Figs. 2B View Fig , 4E View Fig and 5B View Fig ). This trait is unique for the species, and the only other congener that shows a somehow similar morphology is P. diva Gad, 2009 . In this species, the free tips also bend and narrow gradually, but they bend outwards rather than towards each other, and the unfused parts only represents about half of the double organ length, compared to at least 2/ 3 in P. apteryx sp. The unfused tips in P. diva are furthermore densely serrated ( Gad 2009a).
Finally, the arrangement of the double trichoscalids appears to be unique for the new species. Double trichoscalids are present in all species of Nanaloricidae and Pliciloricus , opposite to species of Rugiloricus which only have single trichoscalids ( Fujimoto & Kristensen 2020). However, even though the double trichoscalids in Nanaloricidae are referred to as ‘double’, each trichoscalid in a doublet still attaches to different basal plates. This differs in Pliciloricus where the double trichoscalids attach to the same basal plate ( Neves et al. 2016). In Pliciloricus species known so far, double trichoscalids are either arranged as one trichoscalid on top of the other (e.g. in P. enigmaticus , P. corvus Gad 2005 and P. diva ), or as one trichoscalid appearing as a less developed side-branch of the other (e.g., P shukeri Heiner & Kristensen 2005 ) ( Higgins & Kristensen 1986; Kristensen 1991; Gad 2005c; 2009a; Heiner & Kristensen 2005). In P. apteryx sp. nov. though, the two trichoscalids in the doublet are equally developed and arranged side-by-side with a clear fissure between them ( Fig. 4H View Fig ), rather than being layered as in other congeners. We also consider this arrangement to be unique for the species.
Two additional, not unique but yet uncommon features regard the relative length of the mouth tube and the composition of the midventral plica. Species of Pliciloricus generally have short mouth tubes that are only slightly longer than the corresponding mouth cone. In P. apteryx sp. nov. though, the mouth tube is nearly twice as long as the mouth cone, which is a feature only shared with two other “long-tubed” congeners, P. dubius Higgins & Kristensen, 1986 and P. gracilis Higgins & Kristensen, 1986 . Especially the diamond-shaped appearance of the mouth cone and the long, slender mouth tube in P. gracilis (see Higgins & Kristensen 1986) make the oral parts in this species look very similar to what we see in P. apteryx sp. nov. ( Figs. 2A View Fig , 4D View Fig and 5B View Fig ).
The midventral plica in species of Pliciloricus always differ from other plicae in the lorica. The regular Pliciloricus plicae are separated by primary double ridges, and each plica consists of two, anteriorly arched, smooth longitudinal areas separated medially by a secondary double ridge ( Neves et al., 2016). However, the midventral plica usually differs by being broader than the others, and the occasional presence of additional anterior arches suggests that it sometimes may be formed by fusion of two or more plicae. For instance, the midventral plica of P. cavernicola has six anterior arches, suggesting that it is formed by fusion of three plicae (Heiner et al., 2009). The midventral plica in P. apteryx sp. nov. is rather broad, and the presence of four anterior arches suggests that it is the result of two fused plicae. The midventral plica of P. gracilis is also formed by two plicae, but it is much narrower, with parallel marginal double ridges ( Higgins & Kristensen 1986). The ornamentation of the midventral plica in P. apteryx sp. nov. differs from other species. Whereas the midventral plica of many congeners show a varying number of transverse double ridges connecting with the longitudinal double ridges defining the plica, the two transverse double ridges in P. apteryx sp. nov. do not reach the lateral margins of the midventral plica. Only two other congeners, P. cavernicola and P. dubius , have transverse ridges that do not reach the lateral margins, but the number of ridges differs, with three in P. cavernicola and five in P. dubius ( Higgins & Kristensen 1986; Heiner et al., 2009), opposite to the two ridges in P. apteryx sp. nov.
Other characters that will be of taxonomic significance in future taxonomic and morphological comparison include the shape of the claws in introvert scalid Row 4, the shape and arrangement of basal plates in the neck, and the conspicuous cuticular rosette structures on the ventral side of the lorica. The arrangement and shape of basal plates appear to vary a lot between the congeners, and each species seems to have its own, unique combination. Interestingly, in P. apteryx sp. nov., the shape of the second row basal plates anterior to double trichoscalids show more resemblance with row 1 basal plates in Wataloricus japonicus Fujimoto et al., 2020 than they do with basal plates in any congener (Fujimoto et al. 2020). A midventral rosette has been reported from several Pliciloricus species, but the one in P. apteryx sp. nov. appears much more distinct than in other species. Only P. enigmaticus seems to have a rosette as conspicuous as the one in the new species ( Higgins & Kristensen 1986).
3.1.7. Notes on morphological characters in the Higgins larva
None of the available Higgins larva were containing adult specimens, and we were therefore unable to identify a direct link between larvae and adults. As a consequence, the identity of the larvae is only based on an assumption about conspecifity with P. apteryx sp. nov. However, we consider this assumption to be reasonable well justified, based on the clear congenerity between the larvae and adults, and the fact that no other species of Pliciloricus were found in any of the 31 Bay of Plenty samples collected by Rosli et al. (2016).
The strong contraction of the larval paratypes prevented us from making comparisons with any head structures. However, the thorax and lorica setae, as well as the toes, provide sufficient information to also distinguish Higgins larvae of P. apteryx sp. nov. from other congeners.
All Higgins larvae of Pliciloricus have a pair of anterolateral and anteroventral setae ( Neves et al., 2016). The term ‘seta’ might give the impression of rather simple structures, but they actually show a surprisingly great morphological variation across the known Pliciloricus Higgins larvae. The anterior setae in P. apteryx sp. nov. are short and simple though, which distinguishes them from the short serrated setae in Higgins larvae of P. corvus , P. diva and P. enigmaticus ( Gad 2005c; 2009a; Higgins & Kristensen 1986) and from the longer and branched setae of P. leocaudatus , P. orphanus Higgins & Kristensen, 1986 and P. profundus Higgins & Kristensen, 1986 ( Higgins & Kristensen 1986; Heiner & Kristensen 2005). The anterior setae in the Higgins larva of P. pedicularis is probably those that show closest resemblance with the setae of P. apteryx sp. nov. However, whereas those of the new species appear completely smooth, the setae of P. pedicularis actually show a fine serration near their swollen bases ( Gad 2005b). The lengths of the anterolateral setae are exactly the same in the two species, 24 μm, but the anteroventral setae in P. apteryx sp. nov. are much shorter, only 11 μm, opposite to 24 μm in P. pedicularis .
Also the posterolateral setae, located close to the bases of the larval toes, show some morphological variation. Again, several species show simple setae, but in some species it is also common to find the posterolateral setae attaching on characteristically swollen bases. This swollen bases are present in P. apteryx sp. nov., as well as P. diva , P. leocaudatus , P. pedicularis and P. shukeri ( Gad 2005b; 2009a; Heiner & Kristensen 2005). However, whereas three of the listed species have regular, simple setae, P. apteryx sp. nov. and P. diva show conspicuously swollen setae ( Figs. 2C View Fig , 4I View Fig and 5F View Fig ), which makes their posterolateral setae more than four times as thick as any other setae. In fact, these setae are thicker than the distal ends of the toes, and this distinguishes Higgins larvae of P. apteryx sp. nov. and P. diva from larvae of any other congener.
The combined morphology of the anterior setae and the posterolateral setae in Higgins larvae of P. apteryx sp. nov. makes it possible to distinguish this species from Higgins larvae of any other species.
| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |
Pliciloricus apteryx
| Sørensen, Martin V., Grzelak, Katarzyna, Kristensen, Reinhardt M. & Herranz, Maria 2022 |
Pliciloricus apteryx
| Sørensen & Grzelak & Kristensen & Herranz 2022 |
Pliciloricidae
| Higgins & Kristensen 1986 |
Pliciloricus
| Higgins & Kristensen 1986 |
Loricifera
| Kristensen 1983 |