
Odontesthes perugiae Evermann & Kendall, 1906
|
publication ID |
https://doi.org/10.11646/zootaxa.4250.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:3258EAD0-ED76-4B29-8F7B-2637013FA169 |
|
DOI |
https://doi.org/10.5281/zenodo.6030171 |
|
persistent identifier |
https://treatment.plazi.org/id/03D88787-056D-0630-8FCC-FABBFC3AF6C5 |
|
treatment provided by |
Plazi |
|
scientific name |
Odontesthes perugiae Evermann & Kendall, 1906 |
| status |
|
Odontesthes perugiae Evermann & Kendall, 1906 View in CoL
( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 a, 5a, 6, 7a, 8a, 9a, 10a, 11a)
Odontesthes perugiae Evermann & Kendall, 1906: 94 View in CoL , fig. 3 (original description; type species by original designation; type locality: “Argentina, locality label lost, probably from fresh water”). Fowler, 1943: 326 (listed to Uruguay; material examined from Rio de la Plata, Rio Uruguay at Paysandu, Florida, and Rio Santa Lucia at Canelones). Schultz, 1948: 44 - 45 (data of the holotype). De Buen, 1950: 147 (comparison with O. orientalis View in CoL ). De Buen, 1953: 22, 47 (comparison with O. orientalis View in CoL ). Sverlij et al., 1998: 5, 74 (lower río Uruguay at Salto Grande dam; color photo). Dyer, 1998: 525, 535 (phylogenetic relationships, notes on type locality, senior synonym of Basilichthys microather View in CoL ). Dyer & Gosztonyi, 1999: 214 (phylogenetic relationships). Malabarba & Dyer, 2002: 268, 270, 271 (morphometric and osteological data of the holotype). Nion et al., 2002: 26 (listed to Uruguay). Dyer, 2003: 522 (listed to Argentina, Brazil and Uruguay: “Freshwater drainages of La Plata River”). López et al., 2003: 68 (listed to “río Paraná Medio e Inferior, Delta del Parana, río Uruguay y Río de la Plata”). Liotta, 2005: 554 (listed to Argentina). Dyer, 2006: 70, 73, 74, 81, 84 (referred to the subgenus Odontesthes View in CoL , phylogenetic relationships; identification key). Cuello et al., 2010: 309, 310 (new record to río Negro, Uruguay, and upper río Paraná, Argentina—see comment in “Distribution and type locality notes” section). Loureiro et al., 2013: 102, 110, 112 (conservation status in Uruguay). Serra et al., 2014: 130 (color photo; diagnosis; distribution map in río Negro drainage). Litz & Koerber, 2014: 29 View Cited Treatment (listed to Uruguay). Mirande & Koerber, 2015: 50 View Cited Treatment (listed to Argentina). Almirón et al., 2015: 254 -255 (brief description; photo live specimen; listed to “río Paraná y Río de La Plata”).
Basilichthys microather Marrero Galindez, 1950: 75 -81 View in CoL . Type locality: “Juncales del Río de la Plata y Delta del Paraná” (no types known).
Basilichthys perugiae Lahille, 1929 (new combination). Ringuelet et al., 1967: 475 (material examined, brief description, “Río Paraná inferior; Río de la Plata”).
Odontesthes orientalis de Buen, 1950: 149 View in CoL (“bañado de Legrand, embalse del Río Negro”, Uruguay; holotype and 20 paratypes; holotype illustrated). Ringuelet et al., 1967: 475 (listed as junior synonym of Basilichthys perugiai [sic]. Olazarri et al., 1970: 7 (holotype listed; original number VT 1054 from F. de Buen collection; numbered CI 1808 in the Museo Nacional de Historia Natural de Montevideo; from "Uruguay, Durazno, margen derecho de la boca del arroyo Chileno"). Dyer, 1998: 535 (referred to Odontesthes perugiae View in CoL group). Dyer & Gosztonyi, 1999: 214 (phylogenetic relationships). Malabarba & Dyer, 2002: 268, 270, 271 (morphometric and osteological data of the holotype). Dyer, 2003: 522 (listed to Argentina, Brazil and Uruguay: “La Plata and Uruguay river basins”). Dyer, 2006: 70, 73, 74, 81, 84 (referred to the subgenus Odontesthes View in CoL ; phylogenetic relationships; identification key). Serra et al., 2014: 130 (possibly a junior synonym of Odontesthes perugiae View in CoL ).
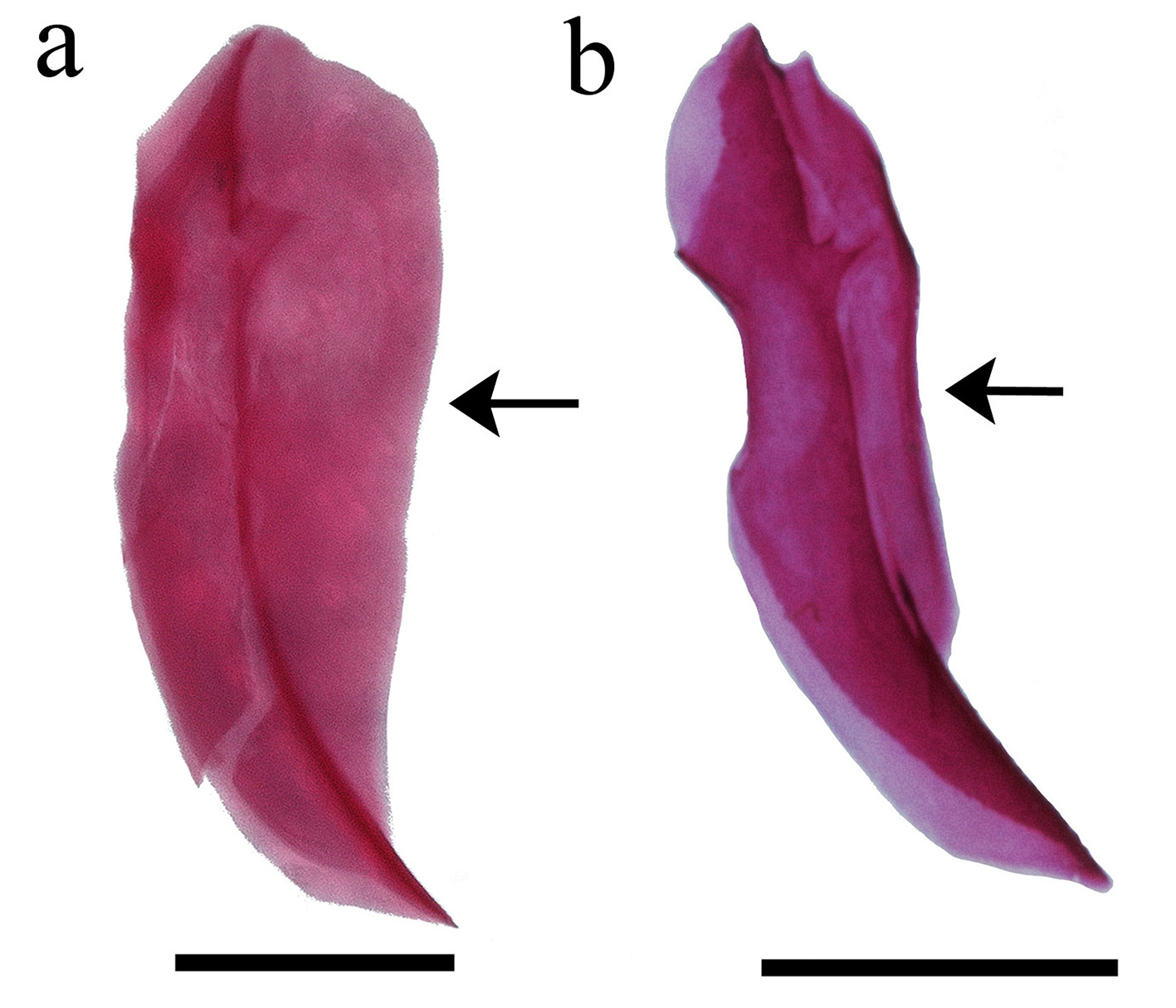
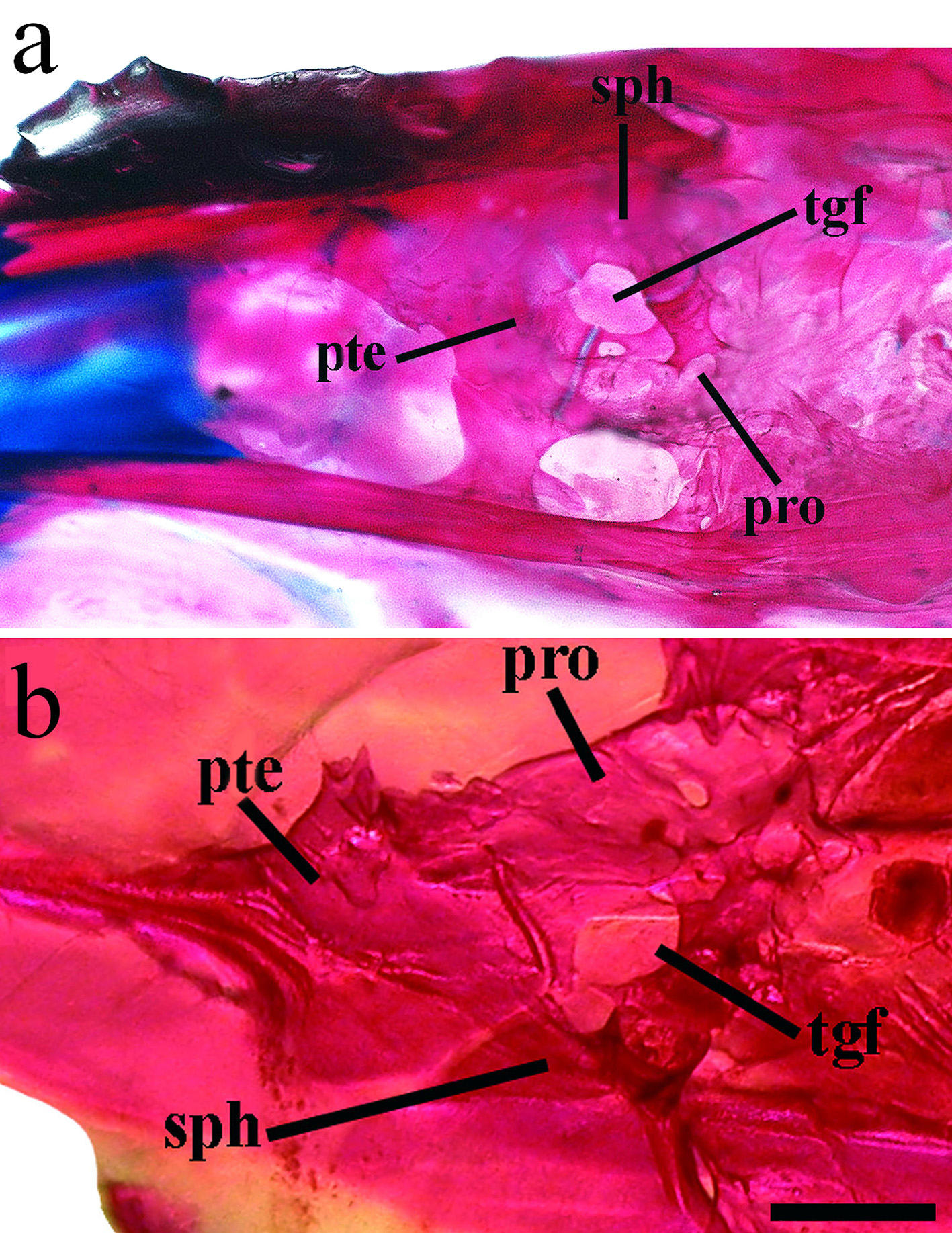
Diagnosis. Odontesthes perugiae is distinguished from congeners, with the exception of the species included in the O. perugiae species-group ( O. bicudo , O. ledae , O. mirinensis , O. piquava and O. yucuman ) by the possession of the following combination of characters: teeth of the outer row on dentary and premaxilla shorter than those in the inner row ( vs. teeth of outer row larger than those of inner row or teeth of outer and inner rows of equal size), origin of dorsal fin positioned over or posterior to anus ( vs. anterior to anus), less than 30 gill rakers on lower branch, and 7 to 9 longitudinal scale rows above left and right lateral bands. Odontesthes perugiae is distinguished from O. ledae , O. mirinensis , O. piquava and O. yucuman by the dermosphenotic with medial flange large, covering its whole extension ( Fig. 4 View FIGURE 4 ; vs. dermosphenotic with medial flange covering only its dorsal portion in O. ledae and O. mirinensis and narrow in O. piquava and O. yucuman ). Odontesthes perugiae is distinguished from O. bicudo by the dermosphenotic sensory canal open ( vs. dermosphenotic sensory canal closed), the basisphenoid with two posterior projections ( vs. basisphenoid with four posterior projections) and the anterior process of the premaxilla shorter than ascendant process ( vs. anterior process of the premaxilla longer than ascendant process). Odontesthes perugiae is further distinguished from O. bicudo , O. ledae and O. piquava by the supraoccipital crest bifid ( vs. single supraoccipital crest). Odontesthes perugiae is further distinguished from O. mirinensis and O. yucuman by the first pleural rib long, attached to ventral postcleithrum ( vs. first pleural rib short, not attached to ventral postcleithrum). Odontesthes perugiae is further distinguished from Odontesthes yucuman by the shorter snout length ( Fig. 5 View FIGURE 5 ; 28.6–37.4 vs. 35.8–39.5% of HL), anterior margin of vomer slightly pointed and lateral condyles distinctly rounded ( Figs. 6 View FIGURE 6 , 7 View FIGURE 7 a; vs. lateral condyles of vomer slightly rounded and anterior margin pointed), the trigeminal foramen enclosed by prootic, sphenotic, and pterosphenoid ( Fig. 8 View FIGURE 8 ; vs. the trigeminal foramen enclosed only by prootic and sphenotic), the palatine with a small rounded dorsal process ( vs. palatine with a distinct rounded dorsal process), the supracleithrum wide curved medially ( Fig. 9 View FIGURE 9 ; vs. supracleithrum narrow), three to five interdorsal pterygiophores ( vs. two interdorsal pterygiophores), the first pterygiophore of second dorsal in two narrow processes ( Fig. 10 View FIGURE 10 ; vs. the first pterygiophore of second dorsal divided in two wide processes), and epural 1 narrow basally ( Fig. 11 View FIGURE 11 ; vs. epural 1 with an anterior process basally).
Description. Morphometric and meristic data in Tables 1—2. Body elongate and slender; dorsal profile slightly convex to nearly straight from snout tip to caudal peduncle. Ventral body profile slightly convex from tip of snout to origin of anal fin; straight and ascending along anal fin and concave near end of caudal peduncle. Caudal peduncle longer than deep.
Head relatively large (16.2—25.1% of SL); dorsal profile straight from snout to posterior tip of supraoccipital; ventral profile of head straight or slightly concave from snout through region below anterior portion of eye. Snout pointed in dorsal view and lateral view ( Fig. 5 View FIGURE 5 a). Mouth terminal, protractile with large gape lying vertically at level of center of eye. Eyes rounded and well developed from lateral view, diameter smaller than snout length.
Pectoral-fin rays i+11—13 (i+11); unbranched and 1st to 4th branched rays longer than remaining rays, its tip not reaching posterior to vertical through pelvic-fin origin. Pelvic-fin rays i+5 (i+5); unbranched ray half-length of first branched ray, not reaching anal-fin origin. Pelvic fins insertion close to each other, with interpelvic membrane uniting them at base or half-length of rays. First dorsal-fin rays iii—vi (iv) with origin at same level or slightly posterior to anus and anterior to anal-fin origin. Second dorsal-fin rays ii+7—9 (ii+7); origin in second half of analfin base. Anal-fin rays ii+12—16 (ii+15) with origin in final insertion of first dorsal fin, distal border usually concave. Caudal-fin rays i+15+i (i+15+i), with scales at least up to half of its length.
Scales large and cycloid with posterior margin smooth; striae forming complete circles. Scales in longitudinal series above lateral band 45—56 (45). Scales in longitudinal series above lateral band before the first dorsal fin 21— 29 (22). Rows of scales between origin of first dorsal fin and origin of anal fin 9—10 (9). Rows of scales between origin of first dorsal fin and posterior insertion of anal fin 8—10 (9). Dorsal scales between posterior insertion of first dorsal fin and origin of second dorsal fin 6—9 (7). Longitudinal scale rows above left and right lateral bands 9— 11 (9). Predorsal scales 22—28. Rows of longitudinal scales in the opercle 6—10. Rows of longitudinal scales below eye 2 (2). Rows of longitudinal scales around caudal peduncle 16—20 (16).
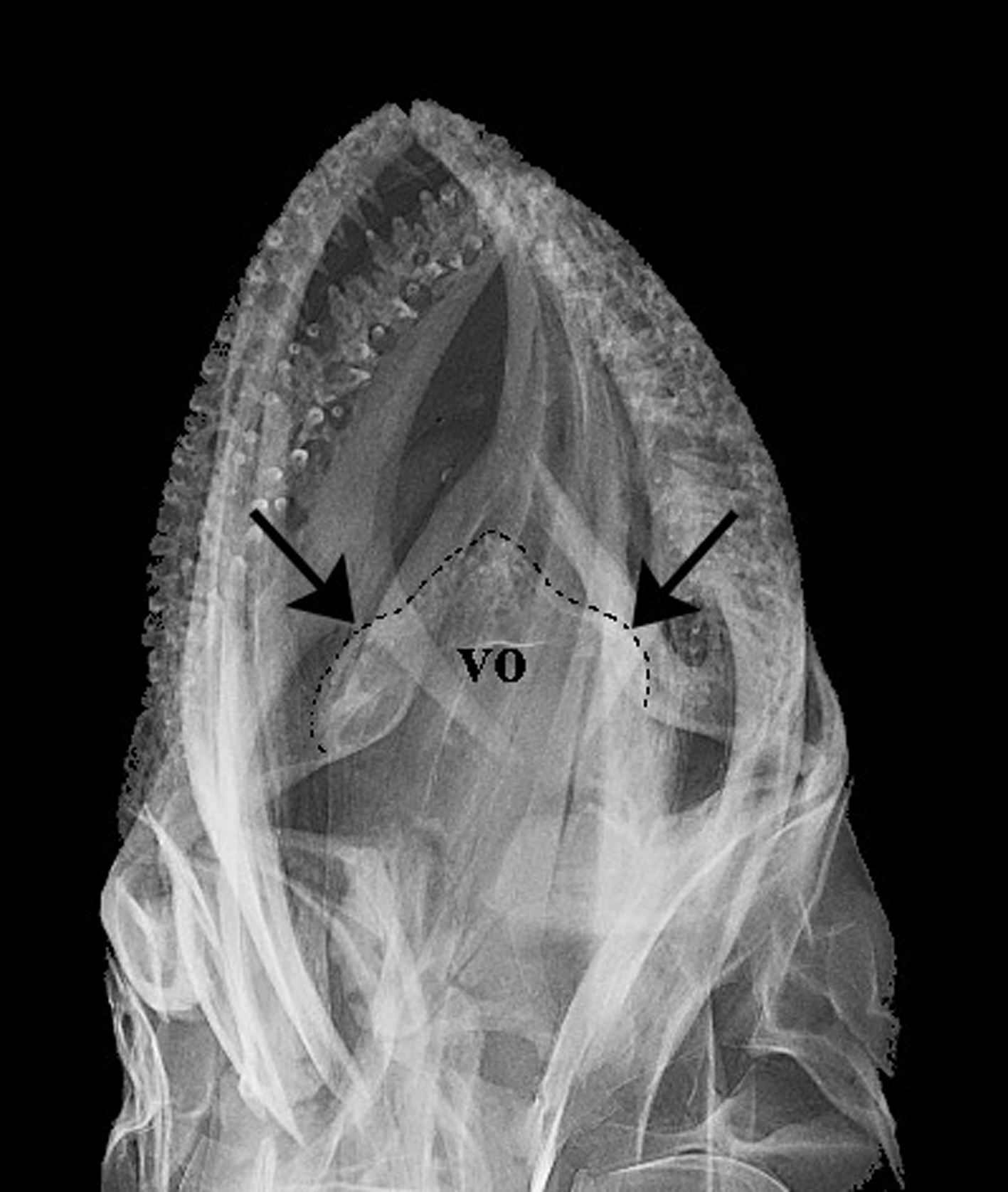
Osteology. Neurocranium. Vomer strongly concave between the anterior tip and lateral condyles; lateral condyles distinctly rounded ( Fig. 6 View FIGURE 6 , 7 View FIGURE 7 a), anterior margin slightly pointed. Three pointed posterior processes, two posterolateral and one central more elongate and extending to parasphenoid. Vomer with three tooth patches with several teeth; tooth patches connected by single row of smaller similar teeth in some specimens. Parasphenoid elongate and narrow with anterior portion wider; posterior portion with narrow lateral wings extending to basioccipital. Basioccipital rectangular, rough and covered dorsally by exoccipital. Exoccipital usually with two foramens and laterally expanded (exoccipital wing); connected dorsally with basioccipital and anteriorly with pterotic. Supraoccipital with forked laminar projection contacting epioccipital dorsally and parietal ventrally. Epioccipital elongate, flattened and with lateral extension, contacting supraoccipital ventrolaterally. Prootic large, trigeminofacial foramen large in contact with prootic, sphenotic and pterosphenoid ( Fig. 8 View FIGURE 8 a). Sphenotic rectangular with anterodorsal process contacting frontal. Dermosphenotic slender with two pores connected dorsally to frontal and medial flange large covering all its extension ( Fig. 4 View FIGURE 4 a). Frontals rectangular with anterior portion rounded, extending more than half skull length. Lateral ethmoid broad, not contacting parasphenoid. Nasal sensory canal detached anteriorly with ventral process pointed and contacting lacrimal subnasal shelf. Lacrimal expanded ventrally, almost contacting infraorbital 2.
Suspensorium. Hyomandibula with dorsal head wide and robust and ventral portion narrow, presenting three articulating surfaces and foramen lodging hyomandibular nerve. Sympletic rectangular and narrow, its anterior portion thinner and posterior portion “v” shaped. Palatine without teeth, with small dorsal process rounded; anterior portion almost contacting quadrate. Ectopterygoid thin, rod-shaped, located between anterior portion of palatine and posterior portion of quadrate. Quadrate contacting anguloarticular anteriorly and endopterygoid dorsally. Endopterygoid well developed with dorsal margin convex contacting quadrate. Metapterygoid elongated and narrow.
Jaws. Maxilla curved, elongate with anterior groove and protruding process posteriorly. Premaxilla with two rows of pointed teeth; teeth of outer row shorter than teeth of inner row; inner row with teeth curved inward; dorsal margin of premaxilla with process ascending into maxilla groove; anterior process smaller than dorsal process.
Rostral cartilage long, almost contacting interpremaxillary ligament. Interpremaxillary ligament narrow, premaxillas close to each other. Dentary with two rows of pointed teeth; teeth of outer row shorter than teeth of inner row; inner row with teeth curved inward; ventral margin slightly convex. Dentary coronoid process broad, higher than anguloarticular coronoid. Meckel’s cartilage elongate and narrow. Anguloarticular coronoid pointed dorsoposteriorly and connected with retroarticular ventroposteriorly. Retroarticular with dorsoposterior portion pointed and connected to quadrate. Coronomeckelian arrow-shaped, located below anguloarticular.
Opercular series. Preopercle curved, with anterior and posterior portions elongated and pointed. Preopercular sensory canal with eight pores; pores 1 and 2 open and pores 3, 4, 5, 6, 7 and 8 enclosed by bone. Interopercle elongated with anterior portion pointed and posterior broad. Subopercle comma-shaped. Opercle broad, with pointed process at superior margin and ventral margin over subopercle.
Branchial basket. Urohyal with one lateral process anteriorly and broad posterior portion. Lower branch gill rakers 19—25; total gill rakers 24—31. Hyoid arch with six branchiostegal rays; four connected in anterior ceratohyal and two connected in posterior ceratohyal. Last three branchiostegal rays widest. Posterior ceratohyal and anterior ceratohyal separated by cartilage, with bony connection in medial portion. Interhyal rectangular and completely ossified connected through cartilage with ceratohyal. Ventral hypohyal rectangular, connected to ceratohyal by bony projections. Basihyal elongate, wider posteriorly. Ceratobranchials 1—4 narrow and long; fifth ceratobranchial enlarged in mid-length with several differently-sized teeth; teeth of outer row longer. Epibranchials elongated and straight with cartilage at tips; epibranchials 1, 2 and 3 with pointed processes at anterior tip, longer in epibranchial 3; epibranchial 4 trapezoidal. Pharyngobranchial 1 small and ossified, pharyngobranchials 2 rectangular, longer than wide with several teeth; pharyngobranchial 3 large and wide with several teeth; pharyngobranchial tooth plate well developed, triangular, with several teeth.
Paired fins. Pectoral girdle with supracleithrum wide, curved medially ( Fig. 9 View FIGURE 9 a). Posttemporal connected ventrally to supracleithrum with two narrow and pointed processes; dorsal process largest. Cleithrum with large dorsomedial wing extending beyond scapular foramen, its ventral portion connected with coracoid. Coracoid partially separated from scapula by cartilage; scapula with elliptical scapular foramen. Radials with ventral and dorsal borders cartilaginous; radials 1 and 2 largest. Extrascapular bone divided in two elements extending over to posttemporal. Pelvic girdle with three pointed processes; dorsal plate well developed, dorsolateral process forming an angle less than 90 degrees with dorsal plate; median process smaller; medial plate reaching anterior tip of dorsal plate.
Medians fins. First dorsal fin with four pterygiophores; first one flat and wider than long. Three to five interdorsal pterygiophores. Second dorsal fin with eight pterygiophores, first one divided in two narrow processes ( Fig. 10 View FIGURE 10 a). Anal fin with 16 pterygiophores, first one widest.
Axial skeleton. Precaudal vertebrae 23—27 (25); caudal vertebrae 21—23 (22); total vertebrae 44—49 (47). Neural and hemal spines inserted anteriorly to vertebrae centrum. First to eighth neural spines extended laterally; fifth to eighth neural spines longer, remaining neural spines elongate and narrow. First haemal spine short and modified forming haemal-arch funnel; haemal-arch funnel beginning between fourth and fifth pterygiophores of anal fin; haemal-arch funnel with lateral expansion.
Caudal complex. Two narrow autogenous epurals ( Fig. 11 View FIGURE 11 a); parhypural near to first hypural. Lower caudal plate with hypurals 1 and 2 fused. Upper caudal plate with uroneural; hypural 3, 4 and 5 autogenous. Hypurapophysis elongate and pointed. Procurrent caudal fin rays 11 (11) dorsally and 12 (12) ventrally.
Color in alcohol ( Figs. 1 View FIGURE 1 and 3 View FIGURE 3 b). Lateral surface of body light yellow with conspicuous longitudinal silvery band in most of specimens extending from pectoral-origin to caudal-fin base, wider below dorsal fins; silvery band with thin black border dorsally. Few specimens with silver band dark gray or completely black after guanine degradation by formalin. Lateral surfaces of body above band presenting small black chromatophores on scales borders. Dorsal surface of body light brown with small black chromatophores in predorsal region, more concentrated on scales borders. Ventral surface of body light brown or beige. Dorsal surface of head and temporal region with dark brown blotches larger than eye size from snout to nape region, two smaller blotches near each nostril. Small black chromatophores on snout, upper and lower lips, anteriormost portion of maxilla, and anteriormost portion of dentary. Fins translucent; pelvic and anal fin without any dark pigmentation. Second dorsal, anal and caudal fins with black chromatophores concentrated basally, becoming inconspicuous towards tips.
Color in life ( Fig. 3 View FIGURE 3 a). Longitudinal lateral band on body bright silvery. Body gray with a silvery tint above lateral band, with lines of chromatophores delineating the scales. Ventral area white silvery, brighter in the belly region, without chromatophores on scales borders. Head dark gray dorsally and silvery in lateral and ventral portions. Snout entirely dark gray. Fins translucent, not differing from preserved specimens.
Sexual dimorphism. No clear sexual dimorphism could be observed in specimens examined.
Size. Maximum length observed 182.2 mm SL. Maximum total length 25 cm according to Sverlij et al. (1998).
Common names. Odontesthes perugiae is commonly known as “peixe-rei” ( Brazil), “flecha de plata”, “matungo”, “juncalero”, “pejerrey”, “pejerrey-juncalero”, “pejerrey-picudo”, “pejerrey de verano”, “pejerrey pigmeo” ( Argentina and Uruguay).
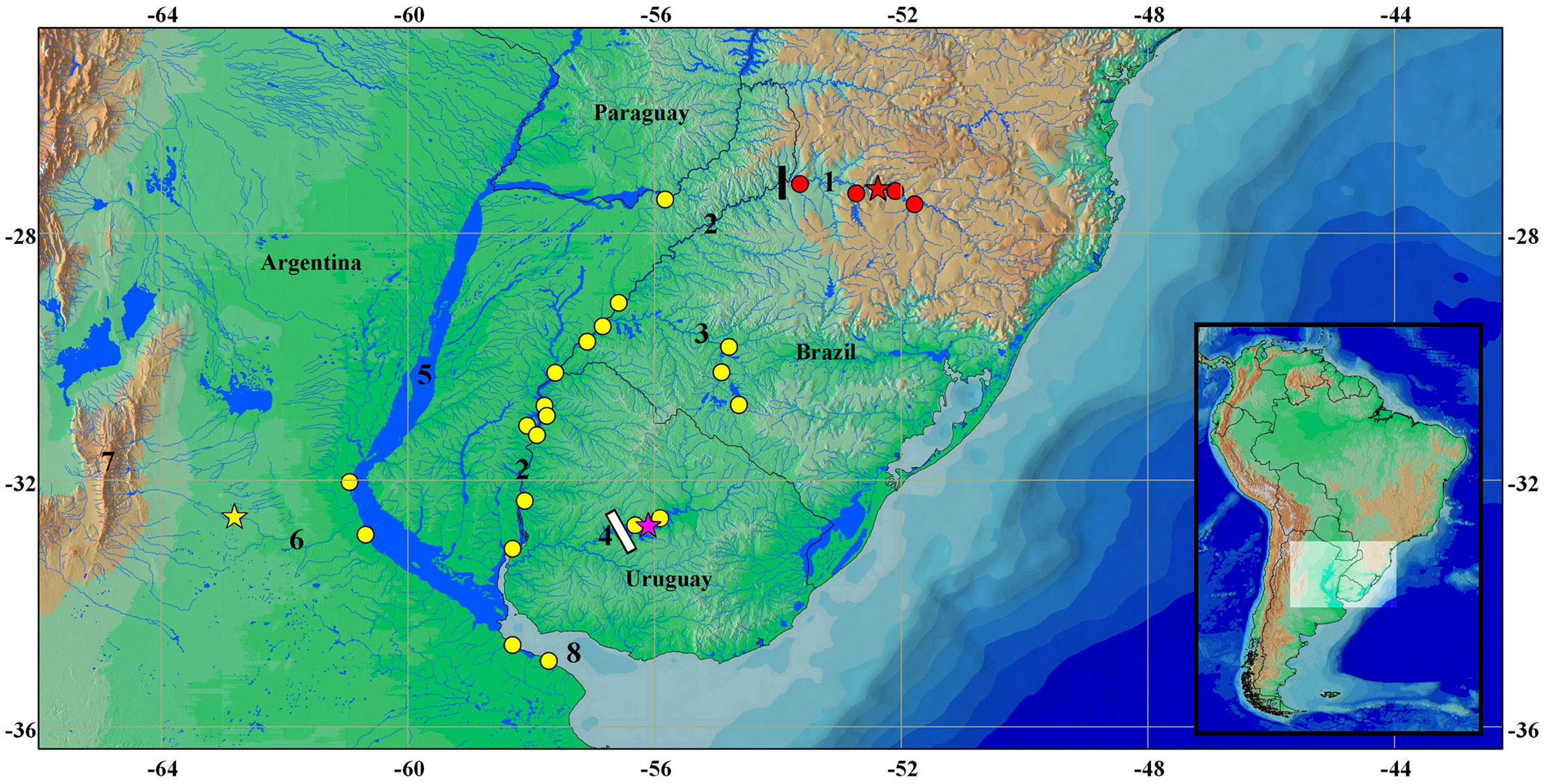
Distribution and type locality notes. Odontesthes perugiae was described based on a single specimen (USNM 55572) collected by John W. Titcomb during an expedition in Argentina between 1903 and 1904. Evermann & Kendall (1906) mentioned the type-locality in the original description as “ Argentina, locality label lost, probably from fresh water”. Evermann & Kendall (1906) mentioned that the freshwater fishes reported by them came from “(…) Rio Primero in the Province of Cordoba, and from the headwaters of the Rio Negro, chiefly Lakes Nahuel Huapi and Traful and tributary or neighboring waters (…).” and that “(…) unfortunately when received some of the labels had become partly effaced, making some of the localities uncertain (…)”. However, some notes by Titcomb cited in Evermann & Kendall (1906) provided valuable information about the freshwater environments explored by him. Among those, the region where the holotype of O. perugiae was most likely collected is described as “streams flowing south from the Sierras in the Province of Buenos Aires ”. These “Sierras” commented by Titcomb likely are the western mountains in the Córdoba Province and their southern tributaries probably refer to those draining to the rio Carcarañá (lower Paraná basin; Fig. 12 View FIGURE 12 ).
Menezes et al. (2003) cited Odontesthes perugiae in a checklist of marine fishes from Brazil as occurring in the western South Atlantic from Brazil to Argentina, but the species is so far know only from freshwaters. Odontesthes perugiae has been cited by several authors to the La Plata basin, all of those records congruent with the known range of the species in the lower rio Paraná, rio Uruguay and rio Negro basins ( Fowler 1943; Ringuelet et al. 1967; Sverlij et al. 2008; Dyer 2003; López et al. 2003; Liotta 2006; Cuello et al. 2010, and Serra et al. 2014). Accordingly, the occurrence of O. perugiae should be restricted to the lower portions of the rio Uruguay (below Salto do Yucumã) and rio Paraná ( Fig. 13 View FIGURE 13 ), corresponding to the lower Uruguay and lower Paraná ecoregions sensu Abell et al. (2008). Recently, Cuello et al. (2010) discussed the distribution of the species, adding records from Corrientes Province in Argentina, referred by the authors as the upper Paraná basin. This locality, however, is located in the lower rio Paraná, as defined in Abell et al. (2008) and other recent papers defining biogeographic regions of freshwater fishes in South America.
Most of the material examined herein was collected in the main channels of the lower rio Paraná, lower rio Uruguay and rio Negro. A few samples were collected in tributaries of the rio Uruguay, in the rio Ibicuí , Brazil, and rio Arapey , Uruguay.
Synonymy accounts. Odontesthes orientalis de Buen 1950 was treated as a junior synonym of Basilichthys perugiai [sic] by Ringuelet et al. (1967). Based solely on the examination of the holotypes of the two nominal species O. orientalis has been repeatedly treated as a valid species ( i.e. Dyer 1998, 2003, 2006; Dyer & Gosztonyi 1999; Malabarba & Dyer 2002), although no diagnostics characters have been presented for its recognition as separate from O. perugiae .
Odontesthes orientalis View in CoL was described based on 21 specimens collected in the rio Negro (rio Uruguay basin), more precisely in a swamp (“bañado de Legrand”) adjacent to the reservoir recognized as Rincón del Bonete ( de Buen 1950) ( Fig. 12 View FIGURE 12 ). The types were originally deposited in the Fernando de Buen private collection (under the number V/T/1054) and, nowadays, only the holotype ( Fig. 13 View FIGURE 13 ) is housed in the MNHM collection ( Olazarri et al. 1970). De Buen (1950) reported the similarity between O. orientalis View in CoL and O. perugiae View in CoL in the original description and provided some diagnostic characters for their recognition: the number of rays in the second dorsal and anal fins, the difference in size of mandibular teeth and the absence of teeth in the vomer. These characters, however, are variable in the samples of Odontesthes perugiae View in CoL analyzed being not useful to diagnose both species. Besides, in contrast to de Buen’s assertion, the holotype of O. orientalis View in CoL also presents teeth on vomer. Three years later, in a revision of silversides from Uruguay, de Buen (1953) redescribed O. orientalis View in CoL and again made a comparison with O. perugiae View in CoL reporting new differences, as such the wider lips in O. orientalis View in CoL and a more posterior position of dorsal fins in O. perugiae View in CoL . The difference in the position of the dorsal fin is notable in the drawings of the holotypes (see Fig. 14 View FIGURE 14 ), but it does not correspond precisely to the condition observed in the holotype of O. perugiae View in CoL , that has most of the first dorsal-fin base located anteriorly to anal-fin base, and not above it as illustrated in the original description ( Fig. 14 View FIGURE 14 ). We extensively compare the types of both species ( Figs. 1 View FIGURE 1 , 13 View FIGURE 13 ) with additional material from the rio Negro (that includes the type locality of O. orientalis View in CoL ), lower rio Paraná (presumably including the type locality of O. perugiae View in CoL ), and lower rio Uruguay and were unable to identify differences justifying the recognition of O. orientalis View in CoL as valid ( Fig. 15 View FIGURE 15 ).
Conservation status. Odontesthes perugiae View in CoL has an extent of occurrence (EOO) larger than 20,000 Km² and is relatively frequently collected, being thus categorized as Least Concern (LC) according to IUCN criteria (IUCN, 2014).
Material examined. USNM 55572 About USNM , holotype of O. perugiae (xr), 110.3 mm SL, J. W. Titcomb col., probably from freshwater, Argentina . MHNM 1808 , holotype of O. orientalis , 104.4 mm SL, reservoir Rincón del Bonete, río Negro, Uruguay . Argentina. CAS 69625, 1 (xr), 120.1 mm SL, Buenos Aires, 34°40’18”S 58°18’08”W GoogleMaps . MACN 5091, 14 , 86.9—125.8 mm SL, río Paraná , Santa Fé, Rosário. MACN 6922, 19 , 92.1—122.2 mm SL, río Colastiné , Santa Fé. MACN 8801, 16 , 87.2—137.5 mm SL, río Uruguay at Embalse La Península , Entre Ríos, Concordia . MCP 33627, 3 , 146.1— 154.7 mm SL, río Paraná , Misiones, Posadas, 27°27’01”S 55°50”W GoogleMaps . UMMZ 218473, 1 (xr), 91 mm SL, playa La Balandra, Buenos Aires, La Plata, 34°55’43”S 57°43’01”W GoogleMaps . UMMZ 231982, 2 (xr), 114.8—131 mm SL, río Gualeguaycito at Salto Grande dam , Entre Ríos, 31°06’34”S 58°04’13”W. Brazil GoogleMaps . All from Rio Grande do Sul State. FMNH 88228, 1 , 79.7 mm SL, rio Uruguai, Uruguaiana . MCP 16198, 16 (1 c&s), 43.8—55.2 mm SL, rio Santa Maria , tributary of rio Uruguai, Rosário do Sul, 30°15’S 54°55’14”W GoogleMaps . MCP 23662, 7 (1 c&s), 42.3-91.4 mm SL, Sanchuri reservoir, Uruguaiana. MZUSP 47689, 4 , 128.6—156.0 mm SL, rio Uruguai, Uruguaiana, 29°46’45”S 57° 07’57”W GoogleMaps . MZUSP 100265, 5 , 103.9— 169.6 mm SL, rio Uruguai, Itaqui.
UFRGS 5205, 1 , 103 mm SL, rio Ibicuí Mirim, Cacequi , 29°50’17”S 54°47’54”W GoogleMaps . UFRGS 8701, 11 , (1 c&s), 50.3—74.0 mm SL, rio Uruguai, Uruguaiana , 29°30’31”S 56°50’45” W GoogleMaps . UFRGS 9348, 1 , 182.1 mm SL, arroio Tacuarembó, Lavras do Sul , 30°46’47”S 54°38’15” W GoogleMaps . UFRGS 10444, 3 (1 xr, ts), 53.5—76.3 mm SL, rio Uruguai, Uruguaiana. 29°30’31”S 56°50’45” W GoogleMaps . Uruguay. ANSP 54097, 3 , 76.2—109.9 mm SL, río Uruguay, Paysandú . MCP 43030, 5 (1 c&s), 94.04—148.5 mm SL, río Uruguay at Salto dam , Salto, Constitución, 31°04’14”S 57°51’24”W. MZUSP 15452, 7 , 100.1— 110 mm SL, río Uruguay GoogleMaps , Salto . UFRGS 7733, 3 , 28.2—125.4 mm SL, río Uruguay at Salto reservoir , Salto, 30°47’20” 57°47’14”W. UFRGS 10443, 1 (1 xr, ts), 106.1 mm SL, río Uruguay , Paysandú, 32°18’48”S 58°06’01” W GoogleMaps . UMMZ 231979, 6 (xr), 46.7—116.7 mm SL, río Uruguay at Salto Grande dam , Salto, 31°15’58”S 57°54’46”W. ZVC-P 5701, 1, 123.2 mm SL, río Uruguay GoogleMaps , Río Negro, 33°06’30”S 58°18’14”W. ZVC-P 8419, 7, 102 mm SL, río Arapey GoogleMaps , Salto, 30°57’16”S 57°45’09”W. ZVC-P 8476, 1, 120.8 mm SL, río Uruguay GoogleMaps , Artigas, Bella Unión, 30°15’35”S 57°36’50”W. ZVCP 6491, 2 , 166.1– 173.2 mm SL GoogleMaps , Tacuarembó. ZVC-P 12081, 3, 172– 205.7 mm SL, arroyo Sauce , Tacuarembó. ZVC-P 12087, 1, 172.5 mm SL, arroyo Las Cañias , Durazno. ZVC-P 12152, 7, 167.1– 179 mm SL, arroyo Malo , Tacuarembó. ZVC-P 12169, 1, 150.7 mm SL, arroyo Las Cañias , Durazno. ZVC-P 12179, 1, 140.1 mm SL, arroyo Laureles , Tacuarembó. ZVC-P 12187, 2 (1 c&s), 156.1-168.7 mm SL, arroyo Tala , Durazno. Exact locality unknown. MCZ 147920, 1 , 141.3 mm SL, rio Uruguay . MACN 4048, 5 , 65.1—133.0 mm SL, rio Uruguay.











| USNM |
Smithsonian Institution, National Museum of Natural History |
| CAS |
California Academy of Sciences |
| MACN |
Museo Argentino de Ciencias Naturales Bernardino Rivadavia |
| MCP |
Pontificia Universidade Catolica do Rio Grande do Sul |
| UMMZ |
University of Michigan, Museum of Zoology |
| FMNH |
Field Museum of Natural History |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
| UFRGS |
Universidade Federale do Rio Grande do Sul |
| ANSP |
Academy of Natural Sciences of Philadelphia |
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Odontesthes perugiae Evermann & Kendall, 1906
| Wingert, Juliana M., Ferrer, Juliano & Malabarba, Luiz R. 2017 |
Basilichthys perugiae
| Ringuelet 1967: 475 |
Basilichthys microather
| Marrero 1950: 75 |
Odontesthes orientalis de Buen, 1950: 149
| Serra 2014: 130 |
| Dyer 2006: 70 |
| Dyer 2003: 522 |
| Malabarba 2002: 268 |
| Dyer 1999: 214 |
| Dyer 1998: 535 |
| Olazarri 1970: 7 |
| Ringuelet 1967: 475 |
| Buen 1950: 149 |
Odontesthes perugiae
| Mirande 2015: 50 |
| Almiron 2015: 254 |
| Serra 2014: 130 |
| Litz 2014: 29 |
| Loureiro 2013: 102 |
| Cuello 2010: 309 |
| Dyer 2006: 70 |
| Liotta 2005: 554 |
| Dyer 2003: 522 |
| Lopez 2003: 68 |
| Malabarba 2002: 268 |
| Nion 2002: 26 |
| Dyer 1999: 214 |
| Sverlij 1998: 5 |
| Dyer 1998: 525 |
| Buen 1953: 22 |
| Buen 1950: 147 |
| Schultz 1948: 44 |
| Fowler 1943: 326 |
| Evermann 1906: 94 |