
Asbestopluma (Asbestopluma) desmophora, Kelly, Michelle & Vacelet, Jean, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.278261 |
|
DOI |
https://doi.org/10.5281/zenodo.6195204 |
|
persistent identifier |
https://treatment.plazi.org/id/03D587B7-FFAD-FFA2-26F7-FA91D5AAB99F |
|
treatment provided by |
Plazi |
|
scientific name |
Asbestopluma (Asbestopluma) desmophora |
| status |
sp. nov. |
Asbestopluma (Asbestopluma) desmophora View in CoL sp. nov.
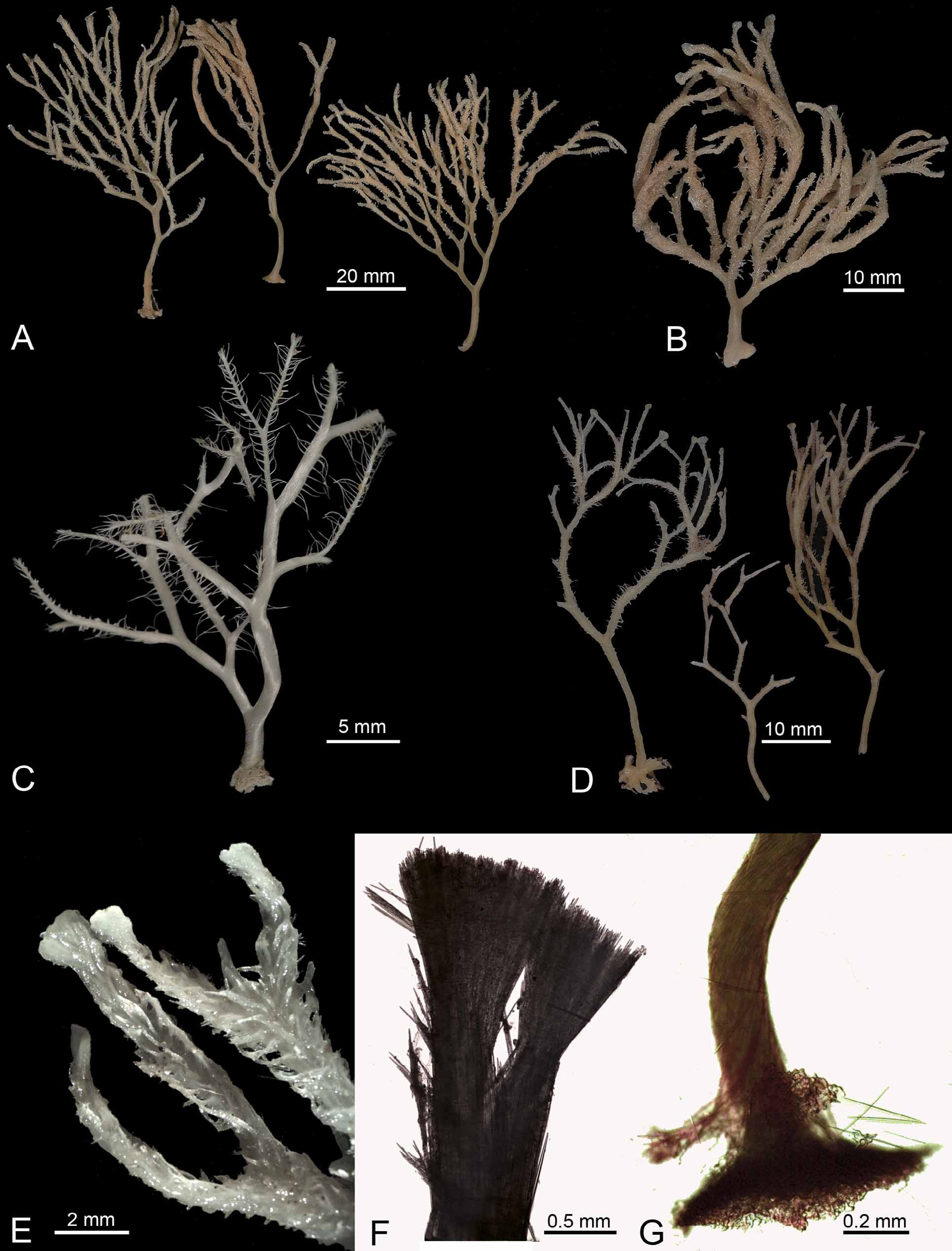
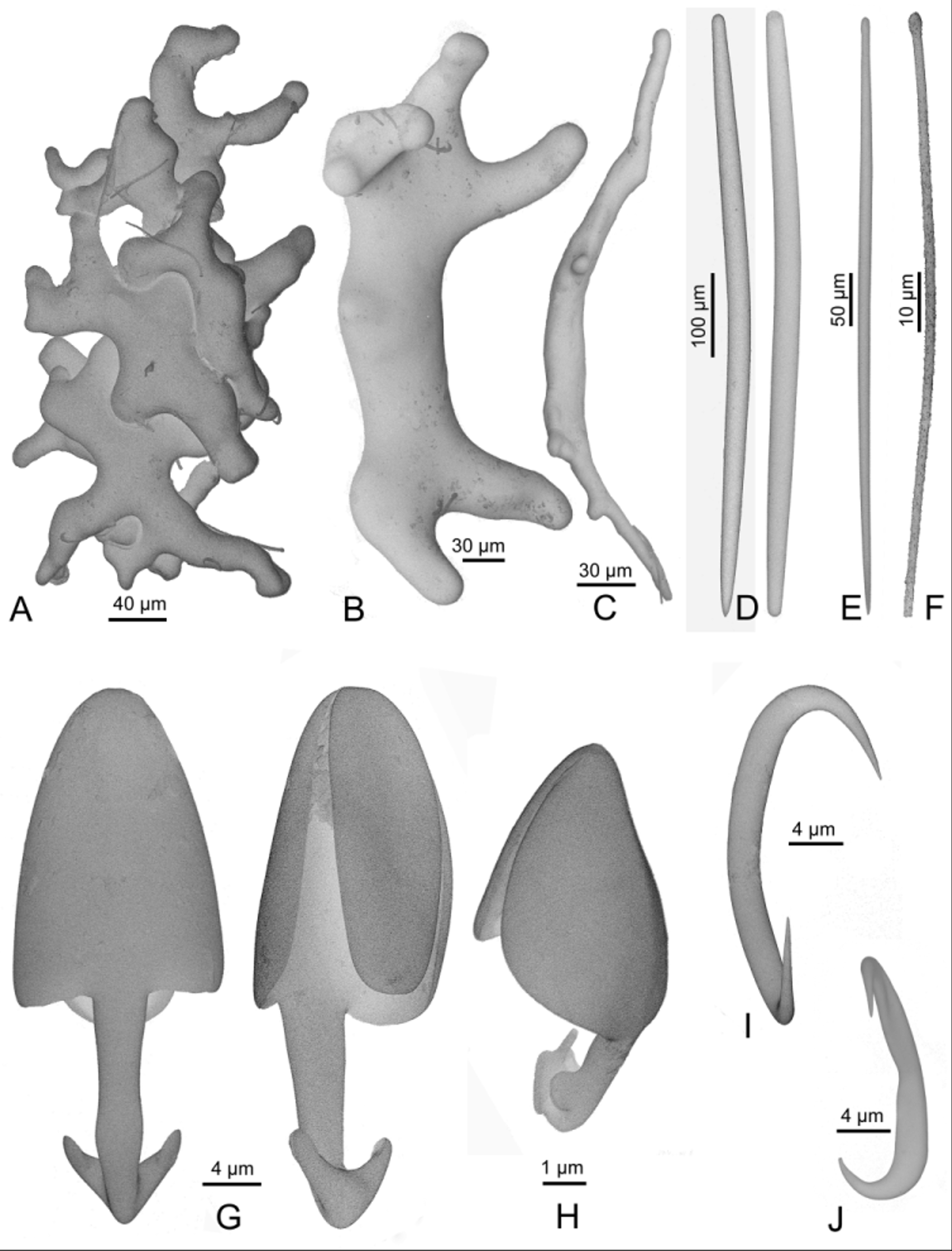
( Figs 5 View FIGURE 5 , 6 View FIGURE 6 ; Tab. 1)
Material examined. Holotype. QM G331844: NIWA Stn TAN0803/102, Seamount 9 Hjort, Macquarie Ridge (Australian EEZ), 56.242°S, 158.462°E, 790–1025 m, 16 Apr 2008, epibenthic sled. Paratypes. QM G331845, QM G331846: NIWA Stn TAN0803/10, Seamount 9 Hjort, Macquarie Ridge, (Australian EEZ), 56.242°S, 158.462°E, 790–1025 m, 16 Apr 2008, epibenthic sled; NIWA 41136: NIWA Stn TAN0803/118, Seamount 10, Macquarie Ridge (International waters), 59.048°S, 158.901°E, 1400–1615 m, 19 Apr 2008, epibenthic sled; NIWA 21350, 21366: NIWA Stn TAN0104/116, Ghoul Seamount, Graveyard Seamounts Complex, northern Chatham Rise ( New Zealand EEZ), 42.798°S, 179.982°E, 1000– 922 m, 17 Apr 2001, epibenthic sled; NIWA 25184: NIWA Stn TAN0604/55, Gothic Hill, Graveyard Seamounts Complex, northern Chatham Rise ( New Zealand EEZ), 42.726°S, 179.897°W, 1030–1140 m, 0 1 Jun 2006, underwater camera; NIWA 21353: NIWA Stn KAH0204/7, Cavalli Seamount, off Northland, North Island ( New Zealand EEZ), 34.119°S, 174.152°E, 800– 670 m, 14 Apr 2002, epibenthic sled; NIWA 35098: NIWA Stn TAN0413/188, Gisborne Knolls, Hikurangi Plateau, off East Cape, North Island ( New Zealand EEZ), 39.019°S, 179.343°E, 2675– 2446 m, 18 Nov 2004, epibenthic sled. Other material. Graveyard Seamounts Complex, northern Chatham Rise ( New Zealand EEZ), Pyre Seamount: NIWA 21351, 21367: NIWA Stn TAN0104/333, 42.718°S, 179.909°W, 1075– 1008 m, 20 Apr 2001, epibenthic sled; Ghoul Seamount: NIWA 62122: NIWA Stn TAN0104/116, 42.798S, 179.982°E, 1000– 922 m, 17 Apr 2001, epibenthic sled; Diabolical Hill: NIWA 25202: NIWA Stn TAN0604/99, 42.790°S, 179.987°W, 890–1160 m, 0 4 Jun 2006, epibenthic sled; Builder ’ s Pencil, Ritchie Ridge, Hikurangi Margin, off East Cape, North Island ( New Zealand EEZ): NIWA 27027: NIWA Stn TAN0616/30, 39.545°S, 178.331°E, 790-815 m, 0 6 Nov 2006, epibenthic sled; Seamount 8, Macquarie Ridge (Australian EEZ): NIWA 40722: NIWA Stn TAN0803/89, 55.381°S, 158.426°E, 504–637 m, 15 Apr 2008, epibenthic sled; NIWA 40796: NIWA Stn TAN0803/91, 55.362°S, 158.428°E, 501–630 m, 15 Apr 2008, epibenthic sled; NIWA 40817: NIWA Stn TAN0803/93, 55.353°S, 158.437°E, 605–709 m, 15 Apr 2008, epibenthic sled.
Type locality. Seamount 9 Hjort, Macquarie Ridge (Australian EEZ).
Description. Numerous specimens and fragments of a fragile, erect, arborescent sponge, up to 80 mm high ( Fig. 5 View FIGURE 5 ). Branches dividing dichotomously, several times in a single plane, with an angle of approximately 45°, the first division occurring generally 4-20 mm above the attachment base. Attachment base hard, slightly enlarged, up to 5 mm in diameter, preserved in a few specimens only ( Fig. 5 View FIGURE 5 A–D, G). First 15 or 20 mm of the axis 1.3-4.2 mm in diameter, devoid of filaments, bare or covered by a feltwork of microtylostrongyles which has been preserved only in a few places. On both sides the branches bear rarely three or four, lateral filaments, which are often reduced to their base but well preserved and up to 4 mm long in numerous specimens, sometimes with brush-like end when well preserved. Consistency of the lower stem and branches rigid, branches are fragile and easily broken. Color, white to cream. No aperture or aquiferous system visible.
Skeleton. Desmas are densely packed and tightly intermingled in the attachment base. In the basal part of the stem, fusiform styles or strongly curved substrongyles form a twisted axis, and the stem is lined by a feltwork of microtylostrongyles preserved in a few places. The axial twist diminishes progressively after the first branch division. Lateral filaments have an axis of mycalostyles, and are frequently anchored through the main axis. Large anisochelae are present only in the terminal zones, where they are attached by their foot, with the large end protruding.
Spicules. Megascleres ( Fig. 6 View FIGURE 6 A–F; Tab. 1), desmas ( Fig 6 View FIGURE 6 A, B), which transition to substrongyles ( Fig. 6 View FIGURE 6 C); 140–450 µm. Mycalostyles ( Fig. 6 View FIGURE 6 D) from the main axis, sometimes substrongyles with a blunt point, varying from fusiform, curved substrongyles located in the basal part of the main axis, to longer, straight, feebly fusiform styles in the upper part of the axis; 400–920 x 12–26 µm, larger in Paratype NIWA 41136 (up to 1500 x 38 µm). Mycalostyles of the filaments with a faintly marked head and an acerate, short point, straight, faintly fusiform, diameter slightly irregular along the shaft ( Fig. 6 View FIGURE 6 E). Intermediates are present with the styles of the main axis in some specimens; 310–600 x 7–12 µm. Microtylostrongyles ( Fig. 6 View FIGURE 6 F) from the basal part of the axis, irregularly curved, minutely spinose, with a round head and an abruptly blunt end; 30–138 µm x 2 –2.5 µm. Microscleres ( Fig. 6 View FIGURE 6 G–J; Tab. 1), anisochelae I ( Fig. 6 View FIGURE 6 G), palmate, head with the two alae entirely linked to the shaft, and a large, ovoid central tooth. Foot with two diverging, short conical alae and no central tooth; 25–40 µm. Anisochelae II ( Fig. 6 View FIGURE 6 H), palmate, abundant, foot with a shaft bearing two round, poorly developed lateral alae, then strongly bent in a central tooth bearing two lateral expansions and ending in a cylindrical point; 7.5–10 µm. Sigmas ( Fig. 6 View FIGURE 6 I), with two acerate ends in different planes, present and uncommon only in some specimens in which they display different size; 10–12 µm in holotype QM G331844, 15– 17 in paratype NIWA 41136, 23–28 µm in NIWA 21353. Sigmancistras ( Fig. 6 View FIGURE 6 J) with two swellings along the shaft and a median depression; 15–27 µm.
Etymology. Named from phor (ancient Greek), meaning to bear (desmas).
Remarks. Asbestopluma (Asbestopluma) desmophora sp. nov. is rather common and quite widely distributed around the New Zealand EEZ, from Cavalli Seamount off North Cape, to Hikurangi Plateau and Margin, off the east coast of the North Island, to the Graveyard Seamounts Complex on the northern edge of the Chatham Rise that extends east from the South Island. It has also been found in the Australia EEZ around Macquarie Island, on Macquarie Ridge that runs southwards from the southwest of New Zealand. It is represented by numerous specimens, some incomplete, and in which, accordingly, some categories of spicules are missing. In many of them, the attachment base and thus, the desmas are absent. The cover of microtylostrongyles has also been lost in many specimens, and generally only the base of the lateral filaments has been preserved. The specimen NIWA 62122, which is devoid of the basal part and which has the filaments relatively well preserved, was mixed with a Cladorhiza sp. from the same location whose fragments are attached to the filaments and to the axis. Anisancorae and large sigmas of this Cladorhiza sp. have been found also in spicule slides of several other specimens. All the incomplete specimens, however, could be identified by the dichotomous branching and by the other spicule characters, especially the shape and size of the anisochelae I and the shape of the anisochelae II. The sigmas and sigmancistras display a variable abundance in the different specimens, and the sigmas seem to be absent in some of them. No significant differences were observed between the specimens from the Chatham Rise and the Macquarie Ridge (Table 1), but neither of the Hikurangi specimens had readily observable desmas in the attachment base.
The species is well characterized in the genus Asbestopluma by the presence of desmas in the attachment base, a character shared among other carnivorous sponges such as Asbestopluma (Helophloeina) stylivarians ( Topsent, 1929) , Euchelipluma arbuscula ( Topsent, 1928) and Esperiopsis desmophora Hooper & Lévi, 1989 . In the new species the desmas appear to derive from curved fusiform styles or substrongyles, as in A. (H.) stylivarians . The species is allocated to the subgenus Asbestopluma rather than to the subgenus Helophloeina , which also may have basal desmas, because Helophloeina is defined by the presence of a cover of microstrongyle and peculiar spearshaped microstyles, which are absent here, and not by the desmas, which are absent in the other known species of the subgenus, A. (H.) formosa Vacelet, 2006 .
It is highly likely that specimens of A. (A.) desmophora sp. nov. devoid of attachment base, and thus without desmas, will be collected. Such specimens are still easy to distinguish from other Asbestopluma spp. by the shape and the characters of the anisochelae. The presence of both sigmas and sigmancistras is also characteristic, but this character is inconstant in our specimens. They can be distinguished from A. (A.) agglutinans Vacelet, 2006 from the East Pacific Rise, which has similar anisochelae, by the external shape, and from A. (H.) formosa from the North Fiji Basin, which also has a dichotomous branching, by the absence of microstrongyles and spear-like microstrongyles, and the presence of sigmancistras. A. (A.) furcata Lundbeck, 1905 from the North Atlantic and Arctic also has a dichotomous branching, but differs by the spicule characteristics.
It is interesting to note that there are several species of carnivorous sponges that have a very similar external morphology. In the present New Zealand collection we have an undescribed Abyssocladia sp. that has exactly the same external characteristics, but which is clearly differentiated by the spicule complement. The two sponges can be distinguished from their external morphological characters only by the brush-like end of the filaments, which are funnel-shaped in the undescribed Abyssocladia sp.. Thus, confidence in the identification of branching species of Cladorhizidae , from in situ images and visual observation, is reduced considerably without the examination of the specimen and its spicules.
| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |