
Paralobesia monotropana ( Heinrich, 1926 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4446.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:843958F9-E4F6-46D3-84B4-108DAA9E6E7A |
|
DOI |
https://doi.org/10.5281/zenodo.5967734 |
|
persistent identifier |
https://treatment.plazi.org/id/03D487C4-516A-8C28-FF51-FBD59EBF260C |
|
treatment provided by |
Plazi |
|
scientific name |
Paralobesia monotropana ( Heinrich, 1926 ) |
| status |
|
Paralobesia monotropana ( Heinrich, 1926) View in CoL
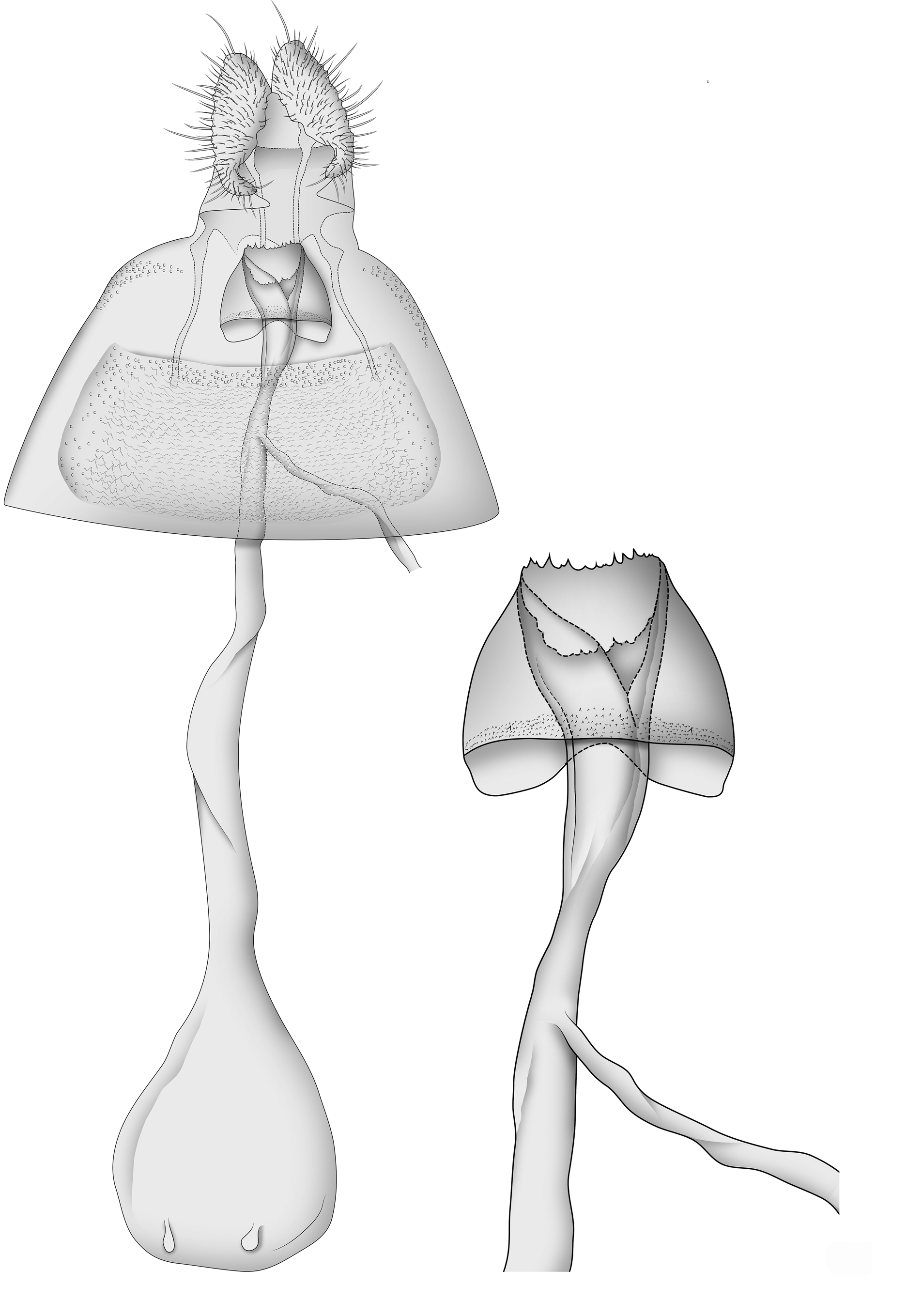
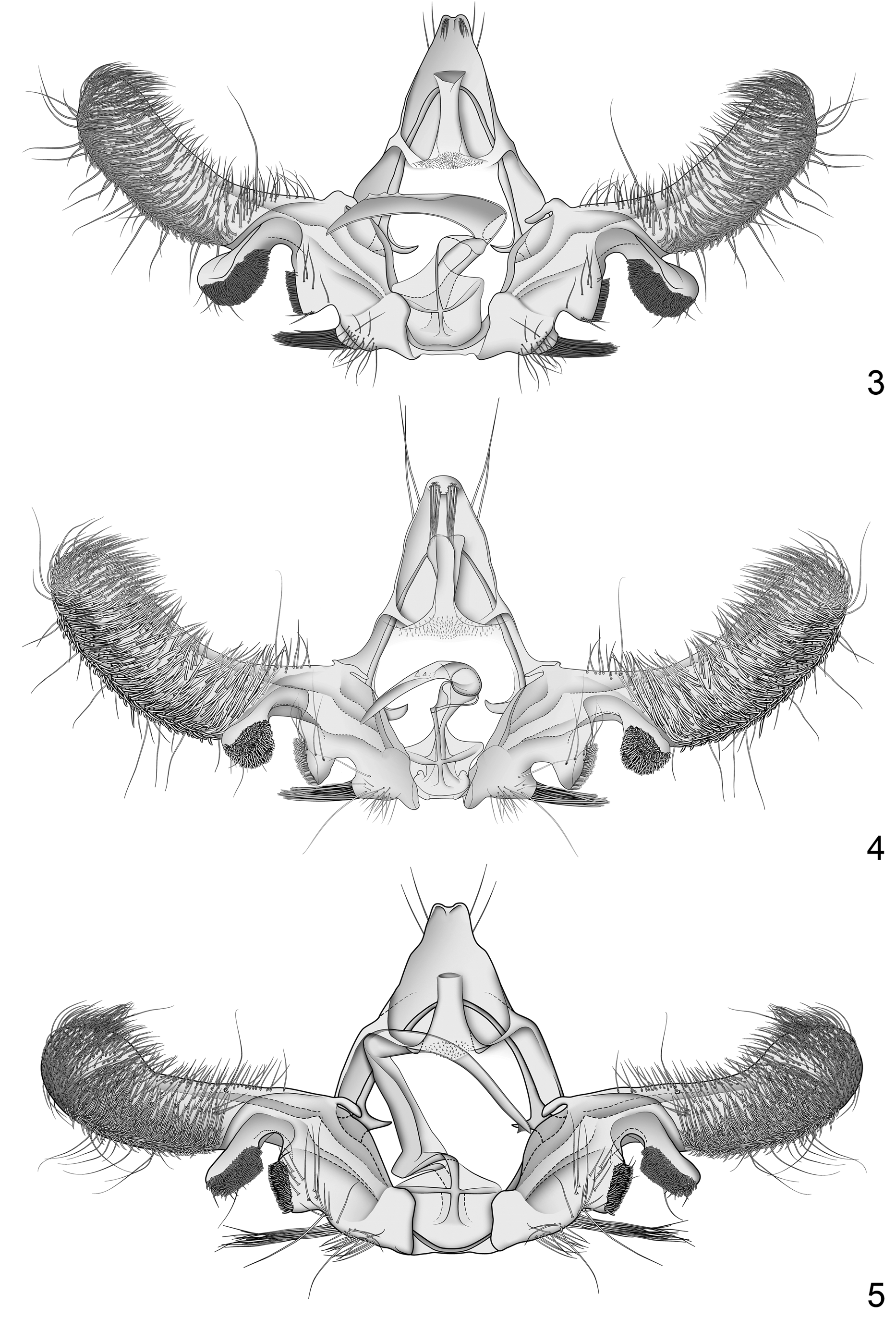
Figs. 3 View FIGURES 3–5 , 6 View FIGURE 6 , 8–12 View FIGURES 8–22
Polychrosis monotropana Heinrich 1926:91 View in CoL ; McDunnough 1939:40; Clarke 1953:229.
Paralobesia monotropana View in CoL ; Obraztsov 1953:92; Brown 2005:472; Gilligan et al. 2008:47.
Endopiza monotropana ; Powell 1983:31.
Diagnosis. Paralobesia monotropana is superficially similar to both P. cypripediana and P. marilynae , but the three species can be separated by features of the male genitalia. In P. monotropana, Spc 1 is ca. 1.5 times as large as Spc2, and the pad of spines extends ca. 0.5 times its length past the ventral margin at the base of the cucullus. In P. cypripediana, Spc 1 is about the same size as Spc2, and the pad of spines extends no more than 0.25 times its length past the ventral margin at the base of the cucullus. In P. marilynae, Spc 1 is about the same size as Spc2, and the pad of spines extends ca. 0.5 times its length past the ventral margin at the base of the cucullus. The emargination between Spc3 and Spc2 is rounded and shallow in P. monotropana , rounded but deeper in P. cypripediana , and shallow but angular in P. marilynae . The phallus of P. monotropana has a series of short wide teeth on the apical 0.33. In P. cypripediana these teeth are smaller and narrower, and extend from a serrated dorsal keel near the middle of the phallus. In P. marilynae there is a single tooth near the apex of the phallus. The setae on posterior surface at the apex of the uncus are shorter than the uncus in P. monotropana , longer than the uncus in P. cypripediana , and absent in P. marilynae . Most other species of Paralobesia that resemble P. monotropana have long (extending past Spc1) setae at Spc3 versus the relatively short setae (not extending past Spc1) in P. monotropana , and a different configuration of teeth on the phallus. In females of P. monotropana , the sterigma is bell shaped and the anterodorsal margin extends past the ventral margin, with a medial depression that often extends posteriorly past the anteroventral margin. That of P. cypripediana is more conical, and the anterior margins are even in length.
Redescription. Male. Head: Vertex pale reddish brown; frons scaling uniformly white; labial palpus pale brown, all segments combined ca. 1.75 times diameter of compound eye, segment II rough scaled, segment III smooth scaled; antenna dark brown. Thorax: Dorsum mottled with reddish-orange and tan scales; posterior crest mottled with dark brown and orange scales; legs pale brown with white annulations on tibia and tarsal segments. Forewing length 4.2–5.2 mm (mean 4.6 mm; n = 9); ground color blue grey, wing markings varying from dark reddish brown to mottled pale brown; costal strigulae pairs 3–9 expressed as pale brown dashes along costa; costal fold absent; patch of elongate scales at base of dorsum; subbasal fascia narrowing from costa to radius, widening from radius to cubitus, narrowing from cubitus to dorsum; median fascia dark brown in costal half with a mix of pale brown in dorsal half, broad from costa to cubitus, distal margin extending towards the termen along the cubitus, and angling back to the dorsum; postmedian fascia divided into two sections, an oval patch at costa and a triangular pretornal patch; postmedian band a large semioval patch extending to termen, usually with notch originating from termen near M3; preterminal fascia a small indistinct patch near apex; fringe scales darkly mottled. Hindwing uniform dark brown with paler scales at base; fringe scales long, dark brown basally, pale brown apically; cubital pecten brown. Abdomen: Pale to dark brown. Genitalia with uncus reduced, weakly bilobed and curved posteriorly, with patch of setae shorter than uncus extending ventrally from apex of each lobe; socius absent; gnathos a weakly sclerotized band, microtrichiate medially, fused with membranous subscaphium; cucullus clavate, stout, costal margin broadly concave, apex narrowly rounded, ventral margin convex with slight medial concavity, ventral half covered in stout spinelike setae, apex and dorsal half covered in fine setae; sacculus with three distinct clusters of spinelike setae, two on padlike lobes proximal to the cucullus and a third on a raised projection at base; Spc1 separated from cucullus by moderate narrow emargination, extending ventrally beyond cucullus ca. 0.5 times its length, Spc1 and Spc2 separated by deep U-shaped emargination, Spc2 0.75 times as large as Spc1, spines on both Spc1 and Spc2 blunt and peglike, Spc2 and Spc3 separated by shallow emargination, Spc3 on a raised lobe, spines on Spc3 stout and spikelike, extending past edge of Spc2. Phallus tapering distally, curved, length ca. 0.66 that of the cucullus, with 3–5 short broad teeth along dorsal margin near apex.
Female. Head: As in male. Thorax: As in male, except forewing length 4.5–5.5 mm (mean 4.9; n = 10). Abdomen: Coloration variable, mostly brown with darker scaling on posterior segments. Genitalia with papillae anales simple, moderately setose; apophyses anteriores ca. 1.5 times as long as apophyses posteriores; sterigma belllike, moderately sclerotized and smooth, with a slight indentation along the dorsal anterior margin, posterior margin serrate; ostium oriented posterodorsally, ductus bursae ca. 2.0 times as long as corpus bursae; ductus seminalis arising in posterior 0.25 of ductus bursae; corpus bursae with paired long, shallow, signum consisting of thickened cells, and two accessory sacs, usually less than 0.1 length of corpus bursae.
Holotype. ♂, “Cincinnati, O., Annette F. Braun, VIII-24-07, on Monotropa uniflora ; 227; Am. Mus. Nat. Hist. Dept. Invert. Zool. No.; ♂ genitalia on slide, CH. 19 May 1922; Polychrosis monotropana Hein. TYPE ” ( AMNH) .
Paratype. USA: Maryland, Cabin John Bridge, R. M. Fouts, “seed capsules of Monotropa ,” 22 Aug 1923 ( 1 ♀, USNM).
Additional specimens examined. CANADA: Ontario: Vineland Station , W. L. Putman, 4 Jul 1942 ( 1 ♂, CNCLEP00105123, slide TOR 5116 About TOR , CNC) ; 20 Jul 1942 (1 ♀, CNCLEP00105122, slide TOR 5117, CNC). Québec: Gatineau Park, Marilyn H. S. Light; pupa from Monotropa uniflora , 16 Sep 2014 ( 1 ♀, CNCLEP 0 0 132701, slide TOR 5115 About TOR , CNC) ; Ramsay Lake , eggs collected 27 Jul 2015, pupated 6 Aug 2015, emerged 22 Aug 2015 ( 1 ♂, CNCLEP00138308, slide HRR 244, CNC) ; Eardley-Masham Road, Trail 56, larvae collected 30 Jul [2016], pupated 6– 8 Aug [2016]; emerged 25 Aug 2016 (1 ♂, CNCLEP00141700, slide HRR 246, CNC); emerged 26 Aug 2016 (1 ♀, CNCLEP00141702, slide HRR 244, CNC); emerged 27 Aug 2016 (1 ♂, CNCLEP 0 0 141703, slide HRR 245, CNC); emerged 28 Aug 2016 (1 ♀, CNCLEP00141704, slide HRR 247, CNC); larva from Rhus typhina , 5 Aug [2016], pupated 7 Aug [2016], emerged 29 Aug 2016 (1 ♂, CNCLEP00141697, slide HRR 248, CNC); USA: Maryland, Washington Co., N.E. Boonsboro Greenbrier St. Park, W. E. Steiner et al., 8–10 Aug 1986 ( 1 ♂, slide HRR 0 37, USNM; 1 ♀, slide HRR 0 34, USNM) ; Wheaton, Homerleigh Rd. , woods, K. Sommerman, 23 Aug 1950, from Monotropa uniflora ( 4 ♂, slides HRR 0 33, HRR 0 35, USNM 124982 About USNM [slide missing], USNM), ( 3 ♀, slides HRR 0 32, USNM 124981 About USNM [slide missing], USNM) .
Distribution and biology. Paralobesia monotropana is recorded from the northeastern U.S. ( Maryland and Ohio) and southeastern Canada ( Ontario and Québec). Rearing records indicate that Monotropa uniflora (Ericaceae) is the primary larval host, although a few specimens have been reared from Rhus typhina (Anacardiaceae) . Observations indicate that this is not an occurrence of accidental oviposition (M. Light, pers. comm.). Females deposit eggs on floral bracts or flower petals, rarely on stems. First instar larvae enter the developing ovary through the pistil. Larvae feed on the interior ovary tissue, but do not feed on seeds; larvae will feed on stem pith if ovary tissue is completely consumed. Collection dates indicate a flight period from early June to late August.
Discussion. Heinrich (1926) listed two female paratypes with identical collection data (USNM), but we were able to locate only one of them.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paralobesia monotropana ( Heinrich, 1926 )
| Royals, Hanna R., Landry, Jean-François & Gilligan, Todd M. 2018 |
Polychrosis monotropana
| Heinrich 1926 :91 |
| McDunnough 1939 :40 |
| Clarke 1953 :229 |
Paralobesia monotropana
| Obraztsov 1953 :92 |
| Brown 2005 :472 |
| Gilligan et al . 2008 :47 |
Endopiza monotropana
| Powell 1983 :31 |