
Mysidium (Orientomysidium), Wittmann & Wirtz, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.495 |
|
publication LSID |
lsid:zoobank.org:pub:65CC1141-E560-4979-97E5-F0701563C84B |
|
DOI |
https://doi.org/10.5281/zenodo.5695543 |
|
persistent identifier |
https://treatment.plazi.org/id/E269F7A6-0187-4AD0-9D64-24C700D5126D |
|
taxon LSID |
lsid:zoobank.org:act:E269F7A6-0187-4AD0-9D64-24C700D5126D |
|
treatment provided by |
Plazi |
|
scientific name |
Mysidium (Orientomysidium) |
| status |
subgen. nov. |
Orientomysidium Wittmann subgen. nov.
urn:lsid:zoobank.org:act:E269F7A6-0187-4AD0-9D64-24C700D5126D
Type species
Diamysis columbiae Zimmer, 1915 , by present designation. This taxon acknowledged in revised combination as Mysidium (Orientomysidium) columbiae (Zimmer 1915) .
Etymology
Noun in nominative singular with neutral ending, formed by amalgamation of the Latin adjective ‘ orientale ’ with the generic name Mysidium , referring to the occurrence on the east coast of America.
Diagnosis
Subdivision of the genus Mysidium Dana, 1852 , characterized by three-segmented carpopropodus of third thoracic endopod; merus of this endopod with serrated setae. Third male pleopod with strong endite at about 50–60% distance from basis of medial margin. Sympod of fourth male pleopod with series of setae on rostral face. Its exopod three-segmented; proximal segment without seta; the two distal segments each with one unbranched modified seta. Telson with terminal cleft.
Distribution
Littoral and sublittoral waters at Atlantic coasts of tropical America (24° N to 8° S), including Gulf of Mexico and Caribbean.
Mysidium (Orientomysidium) columbiae (Zimmer, 1915) subgen. et comb. nov.
Figs 9 View Fig. 9 , 13E–J View Fig. 13
Diamysis columbiae Zimmer, 1915a: 172 –174, figs 23–29 (senior synonym in outdated generic combination).
Diamysis columbiae – Zimmer 1915b: 215 (in synonymy). — Illig 1930: 599 (in synonymy). — Prieto
et al. 2009 (in synonymy).
Mysidia columbiae – Zimmer 1915b: 215, fig. 18 (combination with generic junior homonym); 1918: 26. — Illig 1930: 599 (in key). — W.M. Tattersall 1951: 222 (in homonymy).
Mysidium columbiae View in CoL – W.M. Tattersall 1951: 222, 223 (revised combination). — Davis 1966: figs 1–5. — Brattegard 1969: 86, fig. 27B; 1974a: 69; 1974b: 102, table 1; 1975: 112. — Mauchline 1972: table 3; 1973: tables 2, 4; 1980: 338 (in list). — Băcescu & Ortiz 1984: 22 (in key). — Harrison & Bowman 1987: 678 (in comparison). — Modlin 1987: 115; 1990: figs 1, 3; 1993: 25. — Ortiz & Lalana 1988: 14 (in list); 2018: fig. 10A. — Zoppi de Roa et al. 1989: fig. 2B. — Markham et al. 1990: 411. — Băcescu 1991: 3, fig. 1J. — Escobar-Briones & Soto 1991: table 1. — Ariani et al. 1993: table 1. — Ortiz 2001: 100 (in list). — Price et al. 2002: 44. — Price & Heard 2004: 154. — Sorbe et al. 2007: table I. — Prieto et al. 2009: 33. — Miyashita & Calliari 2014: 9. — Wittmann et al. 2014: 350. — Ortiz et al. 2017a: table 1; 2017b: fig. 3A. — Esteves-Astudillo et al. 2017: 62.
Mysidium colombiae – Harrison & Bowman 1987: 674, 678 (invalid spelling variant). — Price & Heard 2004: fig. 3I; 2009: 938 (in list).
non Mysidium columbiae View in CoL – Brattegard 1969: fig. 27E.
Type material examined
The three vials are from the same sample labelled “Cartagena Kolumbien G. Gagzo leg. 18. IX. 1905 ded. 7. XI. 1906 ”, this material is brittle and delicate, limiting examination. The two specimens from vial 14074 with body parts and appendages sticking together. The respective missing appendages suggest that the lectotype has been partly dissected and figured by Zimmer (1915a: figs 23–29).
Lectotype (by present designation) COLOMBIA • ♂ ad. bl 4.4 mm; registered at the ZMH as “ Diamysis columbiae Zimmer , lectotype by present designation”; 10.37° N, 75.52° W; ZMH K-14073a GoogleMaps
Paralectotypes COLOMBIA • 1 ♀ ad. bl 4.8 mm carrying two postnauplioid larvae; ZMH K-14073b • 1 ♂ ad. bl 4.6 mm, 1 ♀ ad. bl 5.0 mm; ZMH K-14074 .
Other material examined
CURAÇAO • 26 ♀♀ ad bl. 4.4–7.0 mm, 19 ♂♂ ad. bl 3.3–7.4 mm, 11 subad.; #C3; MINGA MYS 431 • 13 ♀♀ ad. bl 5.9–7.0 mm, 7 ♂♂ ad. bl 5.8–7.8 mm, 8 subad.; #C5; NHMW 26479 View Materials .
BONAIRE • 24 ♀♀ ad. bl 4.6–5.3 mm, 22 ♂♂ ad. bl 3.9–6.9 mm, 61 subad.; #B11; ZMH K-55263 • 13 ♀♀ ad. bl 5.0– 7.9 mm, 26 ♂♂ ad. bl 5.7–6.5 mm, 7 subad.; #B12; NHMW 26480 View Materials .
Type locality
Cartagena at the Caribbean coast of Colombia ( Zimmer 1915a). Coordinates estimated by present authors are about 10.37° N, 75.52° W.
Revised definition
All features diagnosed above for the genus Mysidium Dana, 1852 , and its new subgenus Orientomysidium . Cornea globose in lateral view; calotte-shaped in dorsal view, with diameter 1.5–2.0 times as long as terminal segment of antennular trunk. Eyestalks smooth. Rostrum triangular, with concave lateral margins, apically bluntly to acutely pointed, not or weakly extending beyond basis of eyestalks. Anterolateral edges of carapace rounded. Only males with anterior margin of antennular trunk dorsally with rounded, shield-like, mediodistal extension ( Fig. 9A–B View Fig. 9 ) bearing 2–4 setae. The largest seta 0.1–0.2 times extension length. Appendix masculina bilobate, 2.0–3.3 times as long as terminal segment of antennular trunk; all along with brush of long setae. Length of antennal scale 7–8 times maximum width; scale reaching far beyond antennular trunk. Median segment of mandibular palp with setae on both margins. Distally angular hump on outer face of terminal segment of the maxillula. Carpopropodus of thoracic endopods 1–8 with 2, 2, 3, 3–2, 2–3, 2–3, 2–3 or 2–3 segments, respectively. Series of 2–6 serrated setae ( Fig. 9D View Fig. 9 ) along merus of third thoracic endopod. Basal segment occupies half total length of carpopropodus of endopod 3. Pleopod 1 rod-like in both sexes, not considering the weak (sub) median widening at its vestigial endopodal portion ( Fig. 9F View Fig. 9 ). Sympod of male pleopod 4 with strong endite at 2/5 sympod length from basis and with longitudinal series of 3–6 plumose setae on rostral face. Exopod with basal segment occupying 64–72% total length. Endopod reduced to lobe with 27–48% sympod length; with 7–10 barbed setae of various sizes. Endopod of uropods 0.6–0.7 times as long as exopod. Telson subrectangular, length 1.4–1.8 times maximum width near basis. Its lateral margins convex along proximal third; proximal 61–73% of lateral margins smooth, distal portion of each margin with continuous series of 8–12 acute spines. Cleft penetrating 13–15% telson length; cleft separating two broadly rounded, apical lobes; cleft and terminal portion of lobes densely lined in continuous series by a total of 22–30 stout, blunt laminae.
Description of type ( Fig. 9A View Fig. 9 )
All available features fit within the diagnosis. Antennular trunk extends 25–47% its length beyond (artificially aligned) eyes. Sympod of antenna produced into spiniform extension on outer distal corner. Apical segment of maxillary palp about two times as long as basal segment. Carapace posteriorly emarginate, with well-rounded latero-terminal lobes; cervical sulcus strong. Dactylus of thoracic endopods 1–2 with moderately long, strong claw; dactylus 3–7 with long, needle-like claw; dactylus 8 with short seta-like claw. Sympod of fourth male pleopod with 3–4 plumose setae (this range wider in non-types examined) on rostral face plus 1–2 small, barbed setae on inner margin. Pleonites 1–5 are 0.5–0.8, 0.5–0.8, 0.6–0.8, 0.6–0.8 or 0.8–1.1 times the length of pleonite 6, respectively. Scutellum paracaudale triangular with blunt to acute apex; both margins weakly concave to weakly convex. Uropodal endopod 0.9–1.1 times, exopod 1.2–1.5 times as long as pleonite 6. Exopod extends 0.3–0.4 times its length beyond endopod, or 0.7–0.8 times beyond telson; endopod 0.4–0.6 times its length beyond telson. Telson with 0.5–0.7 times as long as exopod of uropods, 0.5–0.9 times endopod or 0.6–0.9 times pleonite 6.
Notes on additional material ( Figs 9B–J View Fig. 9 , 13E–J View Fig. 13 )
Adult females of present material with 4.4–7.9 mm (n = 76) body length, males 3.3–7.8 mm (n = 74). Numbers of serrated setae ( Fig. 9C–D View Fig. 9 ) along the merus of thoracic endopod 3 increase with increasing body size; also numbers of endopods with any setae of that type increase with body size, i.e., pertaining only to thoracopod 3 in smaller individuals and successively to thoracopods 4–7 as individuals become larger. Endopod 8 (when stretched) reaching backwards at most to the end of pleonite 4 and forwards to mandibles; its carpopropodus measures 0.4–0.5 times telson length. Male pleopod 4 comparatively short; exopod reaching at most to basal third of pleonite 6, its subapical seta up to terminal third. Endopod of male pleopod 4 ( Fig. 9H View Fig. 9 ) with the most distal seta 1.4–4.4 times endopod length. Sympod with field of scales on its endite. Statoliths composed of fluorite.
Foregut ( Fig. 13E–G View Fig. 13 ) closely similar to that of M. antillarum sp. nov. ( Fig. 11A–E View Fig. 11 ). As main differences from this species, M. columbiae shows different structure and variable numbers of large spines: on each lateral half there are 2–4 apically pronged spines on posterior part of lateralia, the teeth of these spines with dense sets of secondary denticles ( Fig. 13F View Fig. 13 ); and there are 1–2 larger, apically pronged, serrated spines on dorso-lateral infolding, the latter spines flatter, more clavate, and with more slender teeth on basal ¾ ( Fig. 13G View Fig. 13 ).
Notes on individual development
Nauplioid larvae ( Fig. 13H View Fig. 13 ) with smooth cuticle all around, except for a pair of minute furcal processes and a number of minute setae on the blunt end of the abdomen ( Fig. 13J View Fig. 13 ). Davis (1966: fig. 4) already reported apical, furcal ‘spines’ in nauplioids of M. columbiae from Jamaica. In non-types of the present material, the differentiation of male sexual characteristics is visible in early subadults as small rudiments of penis and appendix masculina and of a bifid fourth pleopod. At this stage the exopod of pleopod 4 is less than 3/2 as long as endopod and the endite of the sympod is missing or indistinct. In the course of further development a distinct endite (representing an important diagnostic character) becomes visible as soon as the exopod exceeds two times the length of the endopod.
Distribution and habitat
From 24° N to 8° S in coastal waters of Bahamas, Caribbean, southern Gulf of Mexico, and Brazil ( Price & Heard 2004; Miyashita & Calliari 2014). The present samples from Curaçao and Bonaire fit within this geographic range. The present material was encountered at a depth of 3–26 m, during daytime in swarms hovering around and between corals. During the night dispersed over the sea floor.
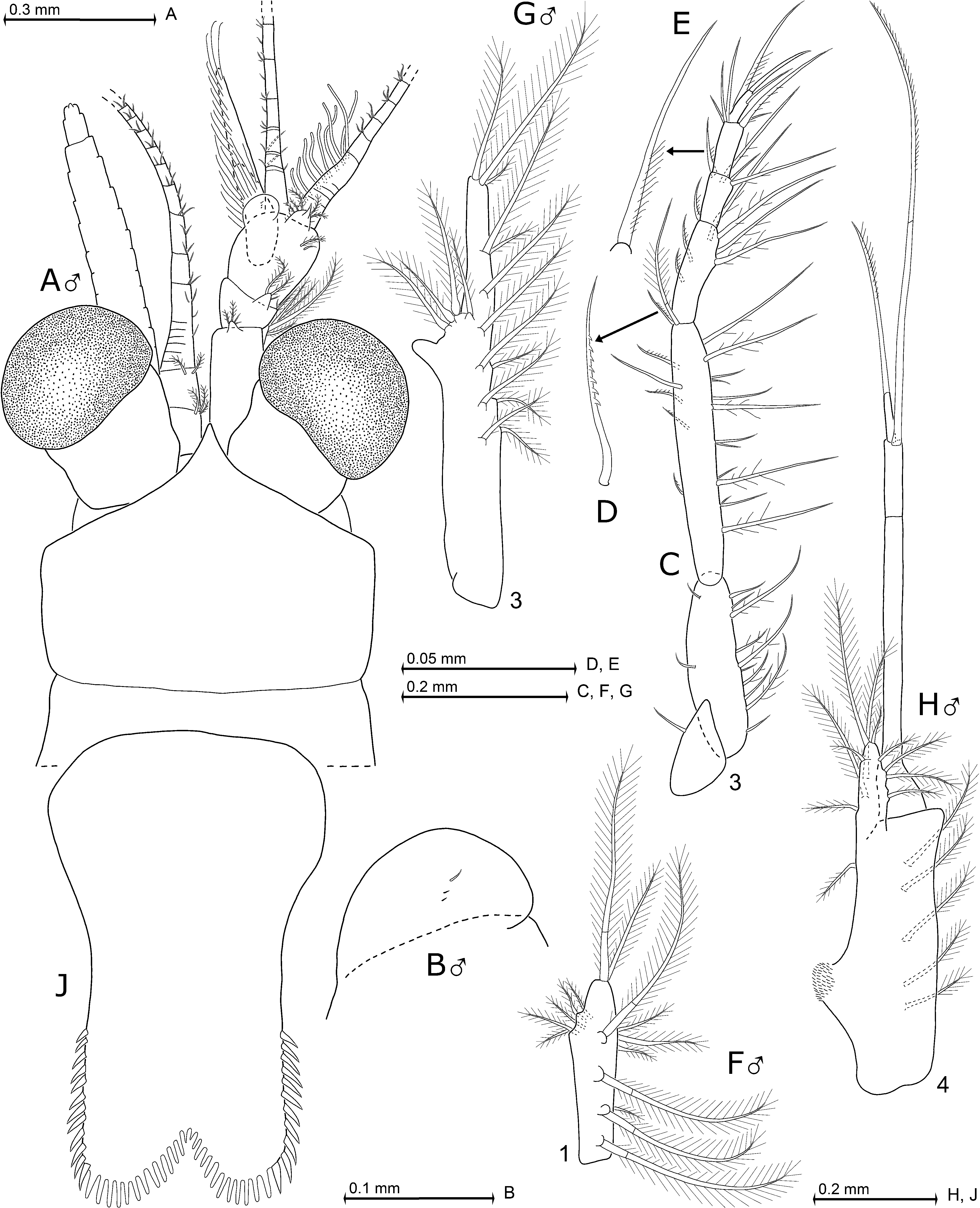


| ZMH |
Zoologisches Museum Hamburg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Mysinae |
|
Genus |
|
|
SubGenus |
Mysidium |
Mysidium (Orientomysidium)
| Wittmann, Karl J. & Wirtz, Peter 2019 |
Mysidium colombiae
| Harrison E. B. & Bowman T. E. 1987: 674 |
Mysidium columbiae
| Esteves-Astudillo M. & Narvaez-Ruiz M. & Ariza-Arredondo L. & Nunez-Penalver J. & Mendoza A. & Marin-Espinoza B. 2017: 62 |
| Miyashita L. K. & Calliari D. 2014: 9 |
| Wittmann K. J. & Ariani A. P. & Lagardere J. - P. 2014: 350 |
| Prieto A. & Alvarez V. & Gonzalez L. 2009: 33 |
| Price W. W. & Heard R. W. 2004: 154 |
| Price W. W. & Heard R. W. & Harris J. T. & McCoy C. M. R. 2002: 44 |
| Ortiz M. 2001: 100 |
| Bacescu M. 1991: 3 |
| Markham J. C. & Donath-Hernandez F. E. & Villalobos-Hiriart J. L. & Diaz-Barriga A. C. 1990: 411 |
| Ortiz M. & Lalana R. 1988: 14 |
| Harrison E. B. & Bowman T. E. 1987: 678 |
| Modlin R. F. 1987: 115 |
| Bacescu M. & Ortiz M. 1984: 22 |
| Brattegard T. 1969: 86 |
| Tattersall W. M. & Tattersall O. S. 1951: 222 |
Diamysis columbiae
| Zimmer C. 1915: 172 |
Diamysis columbiae
| Illig G. 1930: 599 |
| Zimmer C. 1915: 215 |
Mysidia columbiae
| Tattersall W. M. & Tattersall O. S. 1951: 222 |
| Illig G. 1930: 599 |
| Zimmer C. 1915: 215 |