
Phoberogale shareri, Wang & Hunt & Tedford & Lander, 2009
|
publication ID |
https://doi.org/ 10.5252/g2009n4a753 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE660B-C34B-FFAA-FCBD-5B6AFBB88E14 |
|
treatment provided by |
Marcus |
|
scientific name |
Phoberogale shareri |
| status |
sp. nov. |
Phoberogale shareri n. sp. ( Figs 3-5 View FIG View FIG View FIG ; Tables 1; 2)
Cephalogale sp. – Whistler & Lander 2003: 256.
HOLOTYPE. — OCPC 21794 and 21795, rostral part of skull missing tip of inion and much of basicranial region, but with complete upper dentition except right P1, and nearly complete left dentary with c-m2 and m3 alveolus; field number BMM111396.1 View Materials ; collected by Brett M. Malas, Raytheon Infrastructure Services, Inc., December 29, 1995. Although the skull (21794) and dentary (21795) were catalogued with separate numbers, they belong to the same individual, based on their recovery at the same locality and the similar size relationships and stages of wear of their teeth.
ETYMOLOGY. — In honor of Mr Kevin W. Sharer for his leadership as Chairman of the Board of Trustees at the Natural History Museum of Los Angeles County.
TYPE LOCALITY. — OCPC 21794-5 were collected as a result of mitigation activities conducted in support of grading for the Eastern Transportation Corridor ( ETC) highway construction project (Lander 2003). The type locality, OCPC locality 2039, is in the northern ( ETC Windy Ridge) segment of SR 241, about 8.2 km south of its intersection with State Route 91 (Riverside Freeway) and north of its intersection with SR 261 in sec. 23 T.4S. R. 8W. (33°49’34’’N / 117°42’52’’W) in the northern Santa Ana Mountains of Orange County, coastal southern California ( Fig. 1).
DIAGNOSIS. — Phoberogale shareri n. sp. is easily distinguished from “ Cephalogale ” minor Filhol, 1879 [ Filhol 1879] by its much larger size, enlarged P4 and m1, posteriorly located P4 protocone, and differentially enlarged p4 relative to p3. It differs from species of Cephalogale ( C. geoffroyi Jourdan, 1862 , C. ursinus Bonis, 1973 , C. brevirostris ( Croizet, 1836) [ Croizet 1836]) in its larger size and primitively less reduced pre-carnassial premolars and less elongated M2. Within the genus Phoberogale , P. shareri n. sp. is distinguished from European species ( P. bonali Helbing, 1928 , P. depereti Viret, 1929 , P. ginesticus Kuss, 1962 ) by its larger size, and more differentially enlarged p4 over p3. It differs from Pakistani P. bugtiensis in its less prominent notch between anterior base of P4 paracone and protocone, more distinct anterior ridge on P4 paracone, and more mediolaterally shortened M1-2. Species of Phoberogale are distinguished from more-derived Phoberocyon by their smaller size, absence of a premasseteric fossa, and lack of a distinct m2 paraconid.
DESCRIPTION
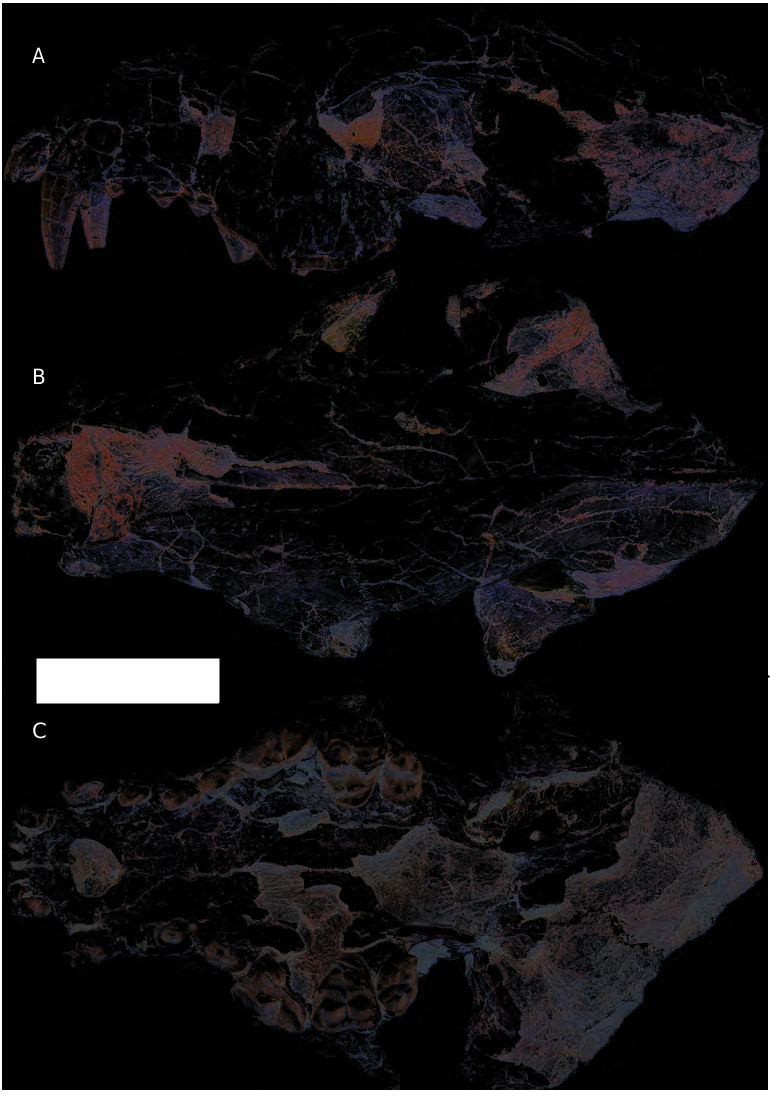
The type and only specimen of Phoberogale shareri n. sp. is preserved in deep purplish mudstones, and suffers from minor to modest distortions and cracks. Its teeth are nearly perfectly preserved. Although the basicranial region in OCPC 21794-5 is missing, it is the only associated skull and dentary in North American Phoberogale materials, and is thus well suited as a holotype. The skull (OCPC 21794) and left dentary (OCPC 21795) are catalogued as separate specimens. Similarities in wear stage on the upper and lower teeth and the preservation of bones suggest that they belong to the same individual.
OCPC 21794-5 belongs to a young adult with permanent dentition fully erupted but showing minimal wear. Th e skull ( Fig. 3 View FIG ) is missing the posterior half of the braincase and the basicranial region, but the remaining parts are more or less intact. Overall, the skull is slightly dorsoventrally crushed resulting in a shallower depth, with the most deformation sustained at the orbital region and the glenoid fossa region.
The nasals sag slightly along the mid line such that the left and right nasals meet each other at almost a right angle. Th e anterior tips of the nasals are missing and the posterior margins extend back behind the anterior rim of the orbit, situated at the same level as the frontal-maxillary suture. The frontal is very broad (widest distance between dorsal rims of the orbits is 59 mm and distance between postorbital processes of the frontals is 55 mm). Th e frontal is slightly elevated behind the postorbital process, indicating a modest development of frontal sinuses. Th e temporal crests behind the postorbital process are not sharply delineated and they converge to a single sagittal crest about 10 mm anterior to the frontal-parietal suture. About 40 mm of the anterior sagittal crest is preserved and it is quite low (~ 4 mm in height).
In lateral view, the infraorbital foramen is at the level of the anterior border of the P4 and the infraorbital canal is relatively long. In anterior view, the foramen is a narrow oval with the long axis oriented vertically. The zygomatic arch is mostly missing on both sides, with only the anterior segment preserved on the right side. The remaining zygomatic arch is quite deep, 25 mm at its anterior base and 14 mm at the broken tip. Bones in the orbital region suffer from extensive damage and the lacrimal area is poorly preserved on both sides. Even on the relatively better preserved left side, individual bones in the orbital mosaic are diffi cult to make out.
In ventral view, the palate is not broadened as in other North American Phoberogale (see Comparison below), with 66.4 mm the maximum distance between the left and right P4s. The incisive foramina are markedly widened and they form a circular opening 12.4 mm in greatest diameter. Part of the palatines, particularly that on the right side near the P4-M1, is missing, and the exact position of the posterior margin is not clear. Judging from the remainder, however, the posterior palatine border seems to stop at a level anterior to the posterior edge of the M2s. Th e basicranium is largely missing, except the left basisphenoid area. An alisphenoid canal is present. The postglenoid fossae have been forced forward by at least 6 mm, resulting in a shortened orbital region, especially on the right side.
The left dentary is largely intact except the tip of the ascending ramus and angular process ( Fig. 4 View FIG ). The horizontal ramus gently tapers toward the anterior end, with the depths and widths below p2, p4, and m2 being 22.2 × 11.8 mm, 22.7 × 11.2 mm, 25.9 × 9.7 mm, respectively.Although the bony surface on the labial side below the m2 is somewhat damaged in an area about 15 mm in width, the remaining bone surrounding this damaged area shows no sign of a premasseteric fossa. Th e bone between the ventral rim of the masseteric fossa and ventral edge of the horizontal ramus is narrow, measuring 8.2 mm in width. Despite the loss of the tip of the ascending ramus, the overall height of the ascending ramus is estimated to be ~ 52 mm. Th e tip of the angular process is missing.
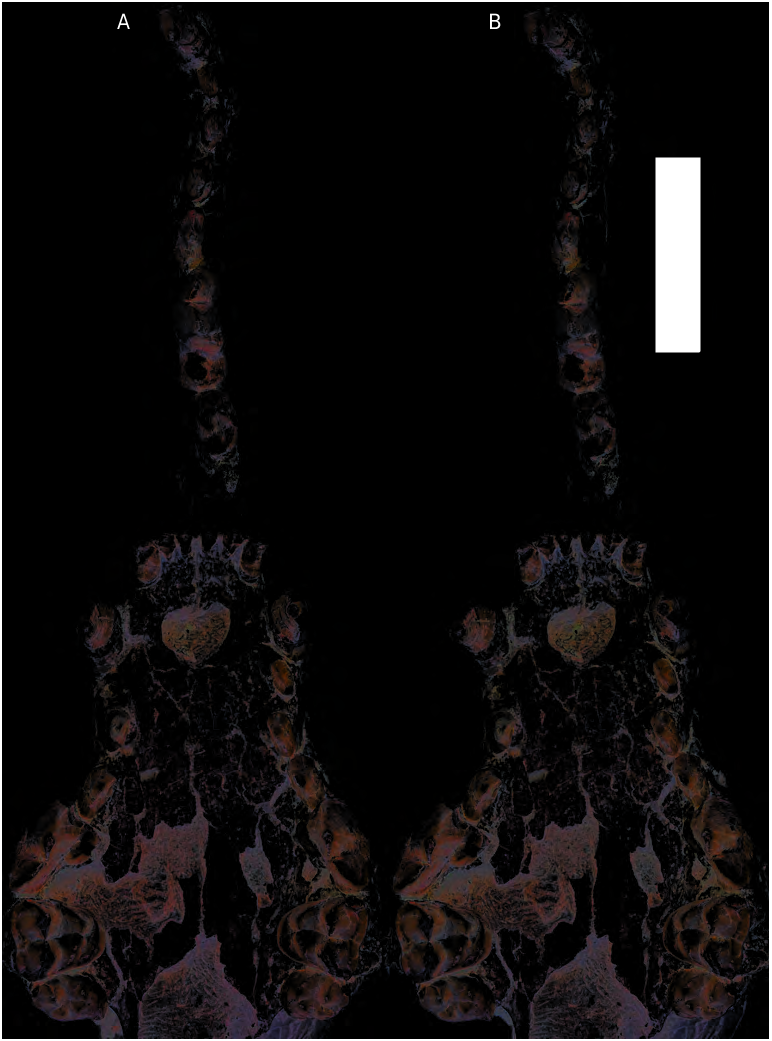
With the exception of the lower incisors, the entire upper and lower dentition is well preserved; only modest wear ensures that the cusp morphology can be described. Th e dental formula is 3142/3143. Upper incisors form a relatively straight dental arcade and they are procumbent. Wear on the tips of the I1-2 obscures their cusp morphology, but they appear to be unicuspid. Th e size increase from I1 to I3 is relatively modest and the I3 is not exceptionally enlarged as seen in some large, hypercarnivorous caniforms. Th e I3 has a medial and lateral ridge, and the medial ridge wraps around lingually to form a basal cingulum. Th e upper canine is conical, oval in cross section, curving backward slightly, and has a total length of ~ 24 mm. An indistinct anterolingual and posterior crest is present along the length of the tooth.
P1-3 are simple, single cusped teeth, and gradually increase in size posteriorly ( Figs 3 View FIG ; 5 View FIG ). All have an anterolingual and a posterior crest. A very indistinct cingulum surrounds the entire P3, and that on the P2 is even more indistinct. P4 is large and robust but not to the extent seen in the large phoberocyonine species from the Nebraska Hemingfordian.Th e paracone has an anterior ridge but lacks a parastyle. Th e protocone is not advanced but is retracted along the lingual face of the P4, hence is posterior to the anterior edge of the paracone, and a slight notch is present between these cusps. The protocone is low and rather flat anteriorly but includes a small sharp, ridge-like cusp toward the posterior end. A very weak cingulum is present on the anterior and labial margin, and that along the lingual margin is more distinct. A distinct carnassial notch is present.
M1 is subquadrate in occlusal outline. The labial cingulum and parastyle are poorly developed. The paracone is taller crowned than the metacone, although wear on these cusps does not allow a more precise description. The protocone is a low, slightly enlarged cusp situated at the lingual termination of the preprotocrista. Th e metaconule forms a low ridge posterior to the protocone, and separated from it by a notch. It is anteriorly expanded at the expense of the postprotocrista. Th e posteriorly directed metaconule ridge adopts a slight labial turn, ending at the posterior margin of the tooth rather than at the base of the metacone. A posterolingual cingulum (hypocone) is moderately developed and wraps anteriorly around the protocone. M2 has an oval outline. Its paracone is much larger than the metacone, and as in M1, the labial cingulum is weak. Th e protocone-metaconule ridge is low and indistinct. Th e lingual cingulum (hypocone) is expanded at its posterolingual corner.
The lower incisors are missing. Th e lower canine is more posteriorly curved than its upper counterpart. The p1-3 are single cusped with an indistinct anterior and posterior ridge ( Figs 4 View FIG ; 5 View FIG ). Size increase from p1-p3 is gradual and slight. The p4 is substantially larger and more robust than p3, and in addition to anterior and posterior ridges, a small posterior accessory cusp is present. The p4 also has a more pronounced cingulum, particularly along the lingual border. The lower carnassial (m1) is long, with a well-defined shearing blade formed by the labial facet of paraconid and protoconid. The protoconid is much taller than the paraconid. The metaconid is low and slightly offset from the protoconid (positioned at the posterolingual aspect of the protoconid).A dominant hypoconid occupies more than two thirds the width of the talonid. A lower entoconid ridge, with less than one third the width of the talonid, encloses a shallow basin. The entoconid crest is not divided into smaller cusps. The m2 protoconid is larger than the metaconid in occlusal view. Wear on these cusps prevents an estimate of their relative height, but it is likely that the protoconid is taller than the metaconid, based on comparisons with other North American Phoberogale specimens. A paraconid is absent, and in its place is a triangular platform enclosed between protoconid and metaconid. As in the m1, the m2 hypoconid is much larger and higher than the entoconid, which is a low crest. An m3 is indicated by a single-rooted alveolus.
GEOLOGY, ASSOCIATED FAUNA, AND AGE
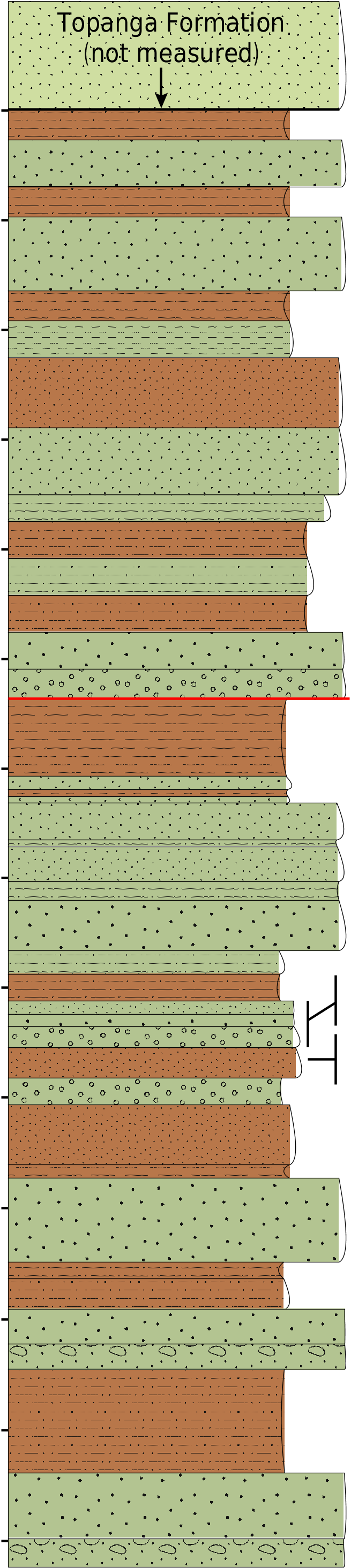
Surficial geologic mapping of the northern Santa Ana Mountains is provided by Morton & Miller (1981) and Schoellhamer et al. (1981). Grading for the northern segment of SR 241 north of Irvine Lake cut through one of the thickest sections of the undifferentiated continental Sespe and marine Vaqueros formations ( Fig. 2 View FIG ). In the northern Santa Ana Mountains, these formations interfinger extensively ( Schoelhamer et al. 1981) and are not easily distinguished or mapped separately. Th e upper or dominantly marine “Vaqueros” lithofacies consists of interbedded reddish, buff, and gray to white sandstones and greenish-gray to white sandstones and siltstones, whereas interbedded reddish, buff, and gray to white sandstones and claystones constitute the lower or continental “Sespe” lithofacies (Lander 2003; Whistler& Lander 2003).Th e formations have a combined maximum thickness of roughly 900 m (2955 ft) ( Schoelhamer et al. 1981). Locally in the ETC Windy Ridge section of SR 41, the exposed, upper portion of the sequence is 317 m (1039 ft) thick and is cut by at least three minor unmapped faults. Th e magnitude and sense of offset along these faults are undocumented. Consequently, the amount of section that has been removed or repeated by faulting is undetermined. Th e base of the section was not exposed by grading. Whistler & Lander (2003) considered the Sespe/Vaqueros Formation in the ETC Windy Ridge section to range in age from late Uintan to early Hemingfordian. The entire section is composed of the “Sespe” lithofacies, which is disconformably overlain by the marine, late Hemingfordian Topanga Formation ( Calvano et al. 2003).
The Phoberogale shareri n. sp. type locality (OCPC locality 2039) was in a 2.7 m (9 ft) thick, finegrained red sandstone 307 m (1006 ft) above the base of the ETC Windy Ridge section, and a greater, but undetermined, distance above the base of the formation ( Fig. 2 View FIG ). This locality is approximately 85 m (280 ft) below the disconformably overlying, marine Topanga Formation of late Hemingfordian age, but the intervening section is cut by the upper of the three documented faults. The holotype (OCPC 21794-5) is preserved in a deep purplish claystone and is the only specimen collected from the site. Stratigraphically correlative sections (ETC Jamboree Road section, Bolero Lookout, Laguna Hills) yielded magnetostratigraphic records variously correlated with C5Cr-C5Er (16.73-19.05 Ma) ( Prothero & Donohoo 2001; Calvano et al. 2008). However, Whistler & Lander (2003) revised the land mammal age assignment for two assemblages in the ETC Jamboree Road section and, consequently, changed the chron assignments to C5Cn-6r (17.28-20.52 Ma).
The upper part of the ETC Windy Ridge section, which contains late Arikareean and early Hemingfordian land mammal assemblages, is above an intraformational unconformity that lies at the base of a conglomeratic interval and separates this part of the section from the lower part ( Calvano et al. 2003). The lower part of the section yielded a late Uintan land mammal assemblage at OCPC locality 2020 ( Calvano et al. 2003; Whistler & Lander 2003).
Whistler & Lander (2003) divided the land mammal assemblages from the upper part of the ETC Windy Ridge section into the upper and lower ETC Windy Ridge local faunas (l.f.s). The early Hemingfordian, upper ETC Windy Ridge l.f. is represented by taxa from OCPC localities 2019 and 2040, whereas the late Arikareean, lower ETC Windy Ridge l.f. spans the “ 290 m of section below locality OCPC 2019” and includes OCPC localities 2016-2018 ( Whistler & Lander 2003). Taxa from the upper ETC Windy Ridge l.f. include Cynelos sp. , Cynarctoides whistleri Wang &Tedford, 2008 , Phlaocyon taylori Hayes, 2000 , Cuyamalagus dawsoni Hutchison in Hutchison & Lindsay, 1974, Nototamias spp. , Miospermophilus spp. , Yatkolamys sp. , and Merychyus elegans Leidy, 1859 , while the lower ETC Windy Ridge l.f. contains Heterosoricidae , Cuyamalagus dawsoni Hutchison in Hutchison & Lindsay 1974, Gripholagomys ? sp. Nototamias spp. , Miospermophilus sp. , Schizodontomys sp. , Leidymys nematodon (Cope, 1879) , Merychyus ? sp.,? Tanymykter sp. ,? Machaeromeryx tragulus Matthew, 1926 , and? Pseudoblastomeryx advena Matthew, 1907 ( Whistler & Lander 2003:table 2; Wang &Tedford 2008). In particular, the murid rodent Yatkolamys sp. from OCPC locality 2019 and the oreodontid artiodactyl Merychyus arenarum or M.elegans from OCPC locality 2062 ( Fig. 2 View FIG ) were considered typical of the early Hemingfordian NALMA and played a key role in separating the upper ETC Windy Ridge l.f. from the lower. Whistler & Lander (2003: 256, fig. 11.4) assigned the Phoberogale shareri n. sp. holotype (OCPC 21794-5) from OCPC locality 2039 to Cephalogale sp. , which at that time they regarded as a member of the upper ETC Windy Ridge l.f. ( Fig. 2 View FIG ).
The Phoberogale shareri n. sp. type locality (OCPC locality 2039) is approximately 56-66 m (185- 215 ft) below OCPC locality 2019, which yielded dominantly early Hemingfordian Yatkolamys sp. and the lowest assemblage assigned to the upper ETC Windy Ridge l.f. by Whistler & Lander (2003), but about 27-39 m (87-127 ft) above OCPC locality 2017 and the highest assemblage referred to the lower ETC Windy Ridge l.f. ( Fig. 2 View FIG ). On the other hand, an undetermined species of Merychyus occurs at OCPC locality 2062, no more than 7.0 m (23 ft) above OCPC locality 2039 (Lander 2003).
Yatkolamys and M. elegans are reported in the early Hemingfordian Runningwater Formation of northwestern Nebraska ( Martin & Corner 1980; Lander 1998: locality CP106). Partly on the basis of the latter two taxa, Whistler & Lander (2003: fig. 11.5) correlated the upper ETC Windy Ridge l.f. and correlative assemblages (including Santiago Canyon Landfill l.f.) in the “Sespe” lithofacies with taxonomically similar assemblages in the Runningwater Formation. In contrast, latest Arikareean Merychyus arenarum , but neither M. elegans nor Yatkolamys , occurs in the Anderson Ranch Formation, which immediately underlies the Runningwater Formation. Found at least 52 m (171 ft) below Yatkolamys , the crushed Merychyus skull (OCPC 22097) from OCPC locality 2062 ( Fig. 2 View FIG ) cannot be assigned confidently to either species. Consequently, the Phoberogale shareri n. sp. holotype from no more than 7 m (23 ft) lower in the section ( Fig. 2 View FIG ) does not occur in association with an age-diagnostic species of this oreodont. Another age-diagnostic taxon in the upper ETC Windy Ridge l.f. is Cynelos sp. , which is based on a partial skull and postcranial skeleton from OCPC locality 2038, no more than 11 m (35 ft) below the Phoberogale shareri n. sp. type locality, OCPC locality 2039 ( Fig. 2 View FIG ). This medium-sized amphicyonid carnivore, presumably an immigrant from Eurasia, is first recorded in North America by a single isolated lower carnassial from the late Arikareean Agate Spring l.f. of northwestern Nebraska, and did not become well represented in the North American fossil record until the early Hemingfordian NALMA of the Great Plains ( Hunt 1972; Tedford & Frailey 1976; Hunt 1998b). Th is skull, based on its proportions and dentition, suggests a latest Arikareean age. Unfortunately, other age-diagnostic taxa, such as the latest Arikareean oreodont Merycochoerus matthewi and early Hemingfordian M. magnus and M. proprius , are not recorded from the undifferentiated Sespe and Vaqueros formations.
Radiometric data from the Sespe Formation support a latest Arikareean age for Phoberogale shareri . n. sp. Along Piuma Road in the central Santa Monica Mountains of Los Angeles County ( Fig. 1), Leidymys nematodon occurs in the lower part of the Piuma Member of the Sespe Formation, whereas Yatkolamys sp. occurs near the top of the upper part of the member ( Fig. 1; Fisk et al. 2001; Lander et al. 2001a; Lander et al. 2001b; Lander 2003; Calvano et al. 2008: fig. 10). Th e two taxa are widely separated stratigraphically, as in the Santa Ana Mountains. Th e Upper Saddle Peak Tuff lies near the base of the upper part of the Piuma Member, roughly 255 m (835 ft) below the Yatkolamys record, and has been determined to be 21.1 ± 0.2 Ma old ( Lander et al. 2003; Calvano et al. 2008: fig. 10). The level that produced the Phoberogale skull probably correlates with one between the tuff and the level that produced Yatkolamys sp.
COMPARISON WITH NORTH AMERICAN FORMS Mainly through intensive collecting by the F:AM and UNSM, such relatively rare carnivorans as Phoberogale were accumulated over time. Although the specimens were collected many decades ago, their existence was announced quite recently and only in the context of North American biochronology, i.e. Cephalogale (now Phoberogale ), along with other immigrants, was used to define the late Arikareean NALMA ( Tedford et al. 1987; 2004).In his summary of North American ursids, Hunt (1998a) briefly discussed the nature of North American Cephalogale . He listed three unnamed species from eight localities: “ Cephalogale new species I” from Standing Rock Quarry, Zia Sand Formation (late Arikareean), Sandoval County, New Mexico; “ Cephalogale new species II” from University Quarry, Upper Harrison Beds (late Arikareean), Sioux County, Nebraska and 16 Mile District, Upper Harrison Beds, Goshen County, Wyoming; and “ Cephalogale new species III” from five different localities in the overlying Runningwater Formation or equivalent beds (early Hemingfordian), Morrill, Sheridan, Dawes, and Box Butte counties, Nebraska. Dental, postcranial and rare cranial fossils constitute the North American material, with the largest sample coming from the early Hemingfordian Bridgeport Quarries.
A comprehensive description of these materials is beyond the scope of this study. Instead, we provide a brief comparison, leaving a full treatment of the remaining North American material for future studies. A decade ago, Hunt (1998a) noted that the largest North American form of Cephalogale (s.l.), his early Hemingfordian species III, exceeded in lower carnassial size (m1 length ~ 23-31 mm) all of the known Eurasiatic forms (~ 12-23 mm). Hunt’s Cephalogale new species I-II were smaller, poorly represented late Arikareean ursids, known by fragmentary teeth only, whereas the larger, early Hemingfordian species III was represented by numerous specimens from the Bridgeport Quarries.
Phoberogale shareri n. sp. is closest in size to Hunt’s (1998b) species II from the late Arikareean of Wyoming (a partial maxilla with M2), as well as small individuals from the Bridgeport Quarries. Compared to the Bridgeport Quarries form, OCPC 21794-95 lacks a parastyle on P4 and consequently lacks a distinct notch between the anterior border of the paracone and the protocone. The California species has a metaconule ridge on M1 closer to the metacone, i.e. directed more labially, in contrast to the more anteroposterior orientation of the ridge in the Bridgeport Quarries species. Th is feature also results in a more developed M1 hypocone (from the posterolingual cingulum) at the expense of the metaconule in the California species as opposed to equal development of metaconule and hypocone in the Bridgeport Quarries carnivore. In addition, the Bridgeport hypodigm displays a distinct paraconule lacking in P. shareri n. sp.
Of the F:AM material, the best preserved specimen is a nearly complete skull and partial skeleton (F:AM 54464) from the “ Cephalogale Ash ” locality 1 mile west of the post office at Marsland, Dawes County, Nebraska. Th is skull shows a rostrum 4% longer than P.shareri n. sp. and a palate 15% broader. This contrast in width is even more pronounced across the zygomatic arches where F:AM 54464 exceeds P. shareri n. sp. by 44%. Similarly, the forehead is broader in F:AM 54464 and the upper canines are more widely spaced ( Table 2). Although the glenoid fossae of OCPC 21794 have been anteriorly displaced, the left alisphenoid canal and area of the orbital fissure are relatively intact. Here the distance between the posterior margin of M2 and the foramen rotundum in F:AM 54464 is 66% greater than in P. shareri n. sp. and this distinction cannot be explained away by mere preservational effects. It is uncertain whether these proportional differences are due to sexual dimorphism, as F:AM 54464 is likely a male with a large upper canine (maximum diameter of 14.6 mm vs. 10.2 mm for OCPC 21794), a large I3 relative to I1-2, and a high sagittal crest.
Other than the above proportional differences between the California and Nebraska skulls, additional characters indicate that they belong to different species. The lateral profile of the sagittal crest in F:AM 54464 is essentially horizontal with a gentle downward curvature near the frontal-parietal suture – the crest is prolonged posteriorly where it forms an overhanging inion. This profile displays a gradual increase in crest depth to the point where the surface of the braincase bends downward. Here the maximum depth of the crest near the inion is ~ 29 mm. Th e crest profile for OCPC 21794, on the other hand, slopes downward in the posterior direction.Th e posterior terminus of the sagittal crest is broken at the point where the braincase surface begins to slope ventrally.Up to this point the sagittal crest shows no sign of increasing depth, which is no greater than 4 mm throughout its preserved length. Hence, a less pronounced inion seems probable in OCPC 21794 and suggests a subtle difference in overall proportions of the temporal muscles and posterior cranium in the two carnivores.

| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.