
Calamagrostis lonana Eggenb. & Leibundg.
|
publication ID |
https://doi.org/ 10.15553/c2023v781a1 |
|
DOI |
https://doi.org/10.5281/zenodo.10722313 |
|
persistent identifier |
https://treatment.plazi.org/id/03CDFC30-D007-FFF5-971E-FCEFFAB5FA54 |
|
treatment provided by |
Felipe |
|
scientific name |
Calamagrostis lonana Eggenb. & Leibundg. |
| status |
|
Calamagrostis lonana Eggenb. & Leibundg. , sp. nov. ( Fig. 2 View Fig , 3 View Fig ).
Holotypus: SWITZERLAND. Valais: commune d’Anniviers , Plateau de Lona , 2588 m, 12.X.2019, Champoud & Eggenberg s.n. ( BERN!; iso-: G [ G00414405 ]!, SION!) .
Calamagrostis lonana Eggenb. & Leibundg. differs from all other Central European Calamagrostis species by its small size: plants are smaller than 20 cm whereas other Calamagrostis species reach (30 –) 50 –150 cm high. Like C. stricta , its callus hairs are distinctly shorter than the lemma and the awn is not protruding from the spikelet but differs by its short panicle with very short branches, short and often glabrous ligule, almost glabrous glumes, and very short anthers.
Perennial, stoloniferous, light to grey-green grass with loose but deep-reaching root system, with extravaginal shoot initiation. Culms solitary, glabrous, (10–)13–18(–22) cm high, erect, hairy towards the inflorescence, with 1 – 3 glabrous nodes; sheaths 1 – 5 cm long, glabrous. Ligules 0.5 – 1 mm long, glabrous abaxially or pubescent towards the tip. Leaf blades 80(–120) × 2–3(–3.5) mm, scabrous adaxially, glabrous abaxially, mostly enrolled. Inflorescences a dense panicle, (2.4–)3.1 –3.7(–4) cm long, gradually spreading, soon contracting after anthesis; panicle branches 1– 3(–4) from main axis, 1 –6 mm long, short ciliate; spikelet stalks 0.5 – 2 mm long. Spikelets 2.5–3.2(–3.6) mm long, lanceolate, 1-flowered, light to grey-green, mostly with a purplish tinge towards the tip. Glumes narrowly lanceolate, long acuminate, the lower 2.6–3.1 mm long, 1-veined, glabrous, the upper 2.7–3.4 mm long, 3-veined, the prominent median vein often ending in an erect tip, fimbriate dorsally, otherwise glabrous; callus short, rounded, covered with hairs 1–1.4 mm long; axillary process tiny, 0.3 – 0.5 mm long, hairy, sometimes absent. Lemmas 2.1–2.4(–2.7) mm long, almost as long as the glumes, 5-veined, pubescent, ragged distally and ciliate like the veins; awn 1.5–1.9 mm long, straight, erect, ciliate, inserted c. 0.6 mm above the lemma base. Paleas 1.3 – 1.6 mm long, delicateskinned, glabrous. Anthers 0.9 – 1.4 mm long, light yellow. Ovaries 0.2 mm long, remaining immature, narrow, with two terminal styles and feathery stigmas c. 1 mm long.
Etymology. – The epithet lonana refers to the name of the Lona plateau in the Pennine Alps (Valais), where the species was found in 2018.
Distribution. – Based on multiple visits of the Lona area, Calamagrostis lonana seems to be restricted to the alluvial plain of the Torrent de Lona and has not been found elsewhere to date. The plain lies at an altitude of 2580 m and extends along the Torrent de Lona, which flows in a west-easterly direction. It is about 720 m long and 200 m wide at its widest point. The distribution area of C. lonana is located within this alluvial plain with a maximum extension of 600 × 150 m. Within this area, several subpopulations of C. lonana could be found. They are very unevenly distributed and dissimilar in density. It cannot be excluded that further localities will be reported from other high altitude alluvial plains in the Pennine Alps.
Ecology. – The large alluvial plain of Lona shows a mosaic of different habitat types which is typical in dynamic alpine alluvial plains ( Fig. 4 View Fig ). In addition to vegetation-free sand and gravel banks, initial alluvial vegetation (Caricion bicolorisatrofuscae), calcareous springs (Cratoneurion) and transitions to fens and transitional bogs (Caricion fuscae, Caricion davallianae, and Caricion lasiocarpae) can be observed. The vegetation classification of the occurrences of Calamagrostis lonana was done by the six releves shown in Table 1. The plot size was 10 square metres for all releves. The abundance was estimated using the Braun-Blanquet cover scale. There is usually an extraordinarily high cover of the moss layer at the sites where C. lonana is growing, a feature that is quite striking and constant. The following moss species were identified: Bryum pseudotriquetrum (Hedw.) G. Gaertn. et al. , Calliergon giganteum (Schimp.) Kindb. , C. richardsonii (Mitt.) Kindb. ex G. Roth , Campylium stellatum (Hedw.) Lange & C.E.O. Jensen , Palustriella falcata (Brid.) Hedenäs [dominant], Philonotis calcarea (Bruch & Schimp.) Schimp. , Polytrichum juniperinum Hedw. , Scorpidium cossonii (Schimp.) Hedenäs , and Warnstorf ia exannulata (Schimp.) Loeske [dominant].
Due to the stratification of the vegetation (dominance of mosses) and the species composition of vascular plants and moss species, the assignment to the habitat type “calcareous spring formation” (Cratoneurion) is most likely. Typical species are Bryum pseudotriquetrum , Palustriella falcata , Philonotis calcarea and the vascular plants Agrostis stolonifera L., Arabis subcoriacea Gren. , and Saxifraga aizoides L. which were often found in the vicinity of the vegetation surveys. The habitat of Calamagrostis lonana can thus be relatively well characterised. It is almost exclusively found in sites with low vascular plant cover and dominance of mosses.
The bedrock across the Lona plateau shows a diversity of different sediment types with both limestone-rich and limestone-poor sediments. In the alluvial plain we measured an average soil pH value of 6.5. The value was confirmed by laboratory measurements of soil samples. The high pH values of the organic soils of the alluvial plain can be explained by
Table 1. – Six vegetation releves showing the co-occurrence of Calamagrostis lonana Eggenb.& Leibundg. with other vascular plants.
Species names according to JUILLERAT et al. (2017). [Abbreviations: imm. = immature; r = very low abundance;
+ = low abundance, cover <5 %; 1 = high abundance, cover <5 %; 2 = cover 5–25 %; 3 = cover 25–50%]
widespread calcareous sediments in the area. The examined soil profiles of Calamagrostis lonana growth sites always show a similar structure: a 10–15 cm thick moss layer ( Fig. 3 View Fig ) is directly followed by a layer of grey alluvial loam more than 50 cm deep, intermixed with dead mosses probably from earlier inundations. The pH value remains unchanged over the entire profile depth (pH 6.5). Roots of C. lonana were found both in the moss layer ( Fig. 3 View Fig ) and in the alluvial loam and can be detected down to a depth of about 40 cm. The root mass of Calamagrostis in a more closely examined soil sample was between 0.11 g (top layer) and 0.02 g or 0.04 g (lower layers) per 100 g of soil material. Whether during inspections at the beginning of August, in September or at the beginning of October the soil was always found to be completely water saturated.
The climate conditions are typical for dry central alpine areas with annual precipitation sum of only approx. 1000 mm (METEOSWISS, 2022). Most of the annual precipitation falls in summer, with a maximum in July. The annual mean temperatures of around 0° C are characteristic of the alpine belt, monthly averages rise to 6° C in summer (METEOSWISS, 2022). However, the basin form of the alluvial plain regularly leads to the formation of cold air depositions, so that temperatures in the alluvial plain are probably well below the mean temperatures of the area. Geomorphological landforms also reveal periglacial conditions across the whole Lona plateau. Mean temperatures are low and frost can occur throughout the year.
Phenology. – Flowering late summer (from August) and fruiting early autumn (from September).
Threats. – It can be assumed that the high plateaus of Lona have been used for grazing by sheep and cattle for a long time (BÄTZING, 2015). The herds can move freely, there is no fencing. From late summer, when the slopes begin to dry out, the herds often gather in the green and fresh alluvial plains and the alluvial plain of the Torrent de Lona is exposed to increased trampling in late August and September. Therefore, the question arises to what extent the population of Calamagrostis lonana might be seriously damaged by trampling and grazing. The trampling impact on the plain is obvious, and tracks remain for a long time. The trampling tracks reveal a lower disturbance in mossy and Cyperaceae -dominated parts of the plain, the sites where C. lonana thrives most. These parts seem to be less attractive for the herds. Judging from the current situation, it does not seem necessary to fence out the alluvial plain. However, long-term monitoring of trampling is recommended.
Notes. – With its short, hairy axillary process, short callus hairs, and long lemmas, Calamagrostis lonana belongs to subgenus Deyeuxia . In central Europe, this subgenus includes the species C. arundinacea , C. varia , and C. stricta . The short, straight awns show a close relationship to C. stricta (GREENE, 1980; ESCALONA, 1988), but it differs from this species in having smaller habit, shorter spikelets with shorter callus hairs, and shorter anthers ( Table 2 View Table 2 ). As in C. arundinacea and C. varia , the spikelets have an axillary process, which distinguishes it from other Central European Calamagrostis species. However, this axillary process is very short in C. stricta (up to 0.5 mm long). Calamagrostis lonana differs from C. arundinacea and C. varia by the short, straight, dorsal awn, which is usually shorter than the lemma. In C. arundinacea and C. varia , the awn is basally inserted, geniculated, and largely exceeding the lemma. Calamagrostis lonana differs from all other Central European species by its small habit, which it also maintains in cultivation conditions. Important morphological characteristics are compared in Table 2 View Table 2 .
The Arctic forms of Calamagrostis stricta show greater morphological plasticity than the Central European taxa, especially C. stricta subsp. groenlandica that shows similarities with the new species. According to literature (see Material and methods), C. stricta subsp. groenlandica is a variable species concerning the habit and the panicle length, displaying panicles as short as (3–)4 cm long. However, the panicles often remain contracted (spreading at anthesis in C. lonana ), the callus hairs are usually more than ½ of lemma length (less than ½ of lemma length in C. lonana ), the ligule is (0.5–)1.5–3(–6) mm long (0.5–1 mm long in C. lonana ), and the glumes are more or less scabrous (glabrous or fimbriate on the keels in C. lonana ).
Based on its morphology and cytology, the taxon found in the Lona alluvial plain belongs to the Calamagrostis stricta aggregate (syn. C. neglecta ). The aggregate is part of the boreal and arctic flora with relict occurrences in Central Europe. Morphologically, the aggregate is very variable and therefore the taxonomy of the whole complex is controversially discussed. Calamagrostis lonana shows the same tetraploid level as all other non-apomictic taxa (2 n = 28, x = 7; ELVEN et al., 2023) and differs from higher-ploidy taxa collected in and around the
[Photo: S. Eggenberg]
Alps. Despite the tiny axillary process, SAARELA et al. (2017) place other tetraploid forms of C. stricta aggregate close to C. canescens (Weber) Roth and C. villosa (Chaix) J.F. Gmel. , i.e., to species that do not possess an axial process. However, the subspecies that often shows small growth, C. stricta subsp. groenlandica , lies outside this clade (SAARELA et al., 2017), which implies a clearer delimitation from C. stricta . Further genetic studies should better resolve the C. stricta aggregate and provide more clues to the taxonomic position of the new species found in the alluvial plain of Lona.
The flower organs develop very late. Anthers and ovaries with feathery stigmas could be observed from the end of September, but no fertile pollen was observed. Conversely, the species shows a strong growth of underground runners. In cultivation, the species shows especially strong vegetative growth and forms dense, vigorous mats within one growing season.
The 2 n -genome size estimates obtained by flow cytometry varied between 7.33 pg and 8.10 pg of DNA for samples of Calamagrostis lonana , perfectly matching estimates for tetraploid Calamagrostis taxa (2 n = 28) published in the literature (SMARDA et al., 2019; ZONNEFELD, 2019). In contrast, all other samples of presumably C. stricta collected across nearby locations in France and Germany presented genome size estimates ranging from 14.39 pg to 16.50 pg, rather matching decaploid taxa of Calamagrostis with 2 n = c. 70 chromosomes.
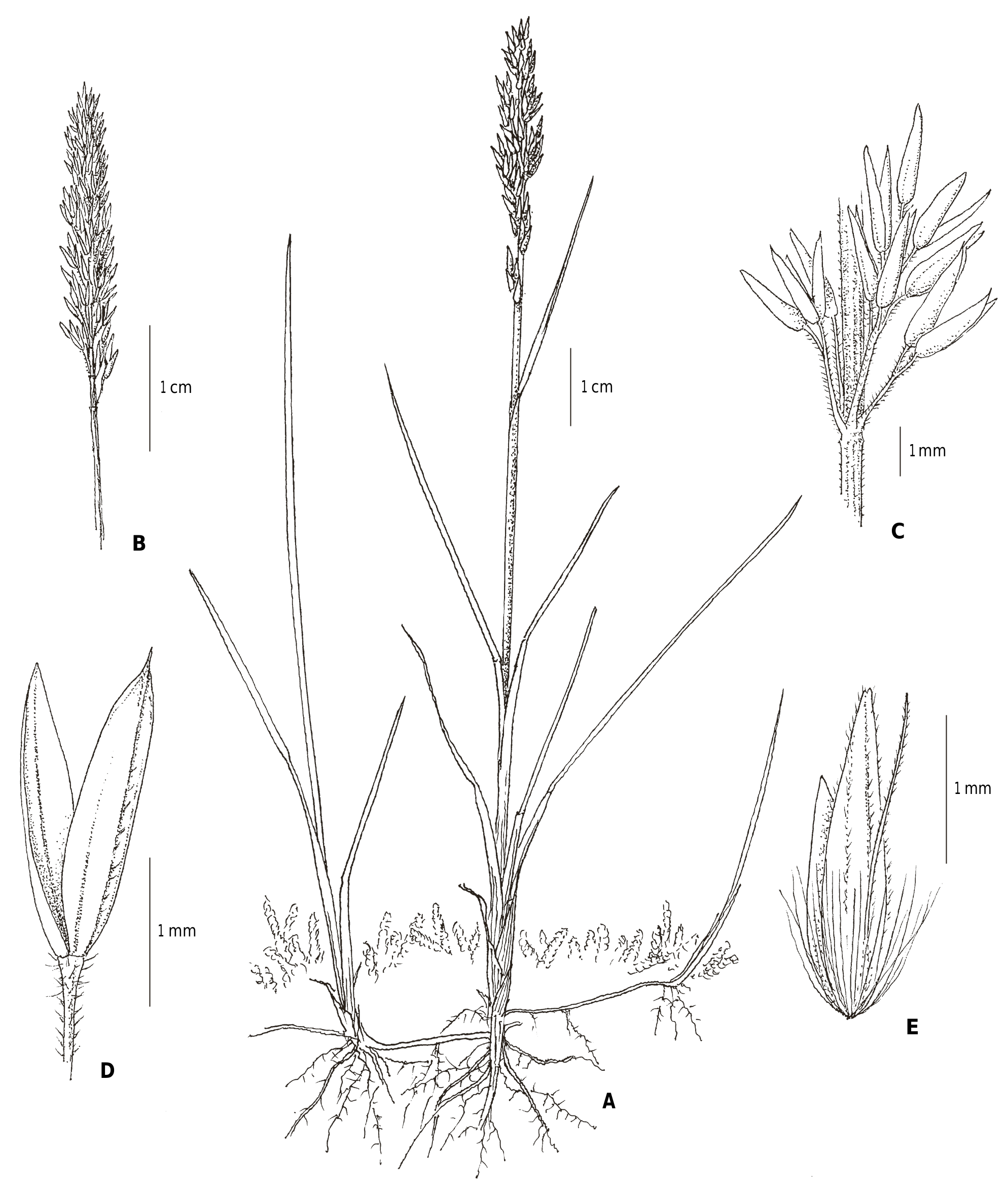


Table 1. – Six vegetation releves showing the co-occurrence of Calamagrostis lonana Eggenb.& Leibundg. with other vascular plants. Species names according to JUILLERAT et al. (2017). [Abbreviations: imm. = immature; r = very low abundance; + = low abundance, cover <5 %; 1 = high abundance, cover <5 %; 2 = cover 5–25 %; 3 = cover 25–50%]
| Plot ID | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Total cover | 98 % | 95% | 95% | 90% | 100% | 95 % |
| Cover of herb layer | 10 % | 15 % | 20% | 15 % | 30% | 15 % |
| Cover of moss layer | 95% | 95% | 92 % | 85% | 95% | 95 % |
| Calamagrostis lonana | + | 1 | 2 | 2 | + | 2 |
| Eriophorum angustifolium | 1 | + | + | + | r | 1 |
| Eriophorum scheuchzeri | 2 | 1 | ||||
| Eleocharis quinqueflora | 1 | 2 | ||||
| Carex rostrata | 1 | 3 | ||||
| Equisetum variegatum | + | |||||
| Juncus triglumis | 1 | + | ||||
| Polygonum viviparum | r | |||||
| Salix hastata (imm.) | r |
Table 2. – Comparison of morphological characters between Calamagrostis lonana Eggenb.& Leibundg. (own measurements) and the Central European C. stricta (Timm) Koeler (measurements retrieved from HESS et al., 1967–1972;QUINGER, 1987; CONERT, 1998; MÜLLER & al., 2021; PAROLLY & ROHWER, 2016).
| C. lonana | C. stricta | ||
|---|---|---|---|
| Habit | Culm length [cm] | (10–)13–18(–22) | 30–100(–120) |
| Size [mm] | 80(–120) × 2–3(–3.5) | up to 600 × (1.5–)2–4(–5) | |
| Leaves | Colour and form Ligule length [mm] | light green to greyish green, mostly enrolled 0.5–1 | fresh to greyish green, flat or rolled 2–3 |
| Ligule indument (abaxial side) | glabrous | hairy | |
| Length [cm] | (2.4–)3.1–3.7(–4) | 8–15(–20) | |
| Panicles | Shape Colour | contracted but spread at anthesis grey to lilac | contracted, rarely spread at anthesis grey to pale lilac |
| Branche length [mm] | 1–6 | 15–60 | |
| Length [mm] | 2.5–3.2(–3.6) | 3–4(–4.5) | |
| Glume length [mm] | 2.6–3.1 | 3–4(–4.5) | |
| Callus hair length [mm] | 1–1.4, <½ of lemma length | 1.5–2.5, ½ to ¾ of lemma length | |
| Spikelets | Axillary process length [mm] Lemma length [mm] | 0.3–0.5 2.1–2.4(–2.7) | 0.8–1 2.6–3.5 |
| Awn length [mm] | 1.5–1.9 | 2–3 | |
| Palea length [mm] | 1.3–1.6 | 1.8–2.3 | |
| Anther length [mm] | 0.9–1.4 | 2–2.5 | |
| Cytology | Chromosome number | 2 n = 28 | 2 n = c. 70 |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |