
Paradeontacylix godfreyi, Hutson & Whittington, 2006
|
publication ID |
https://doi.org/10.11646/zootaxa.1151.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:8E79FB4C-7C7A-4DBF-B799-6CDD9E8D9CF3 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB87F3-4430-787B-FEA3-3DE2FC4BFE35 |
|
treatment provided by |
Felipe |
|
scientific name |
Paradeontacylix godfreyi |
| status |
sp. nov. |
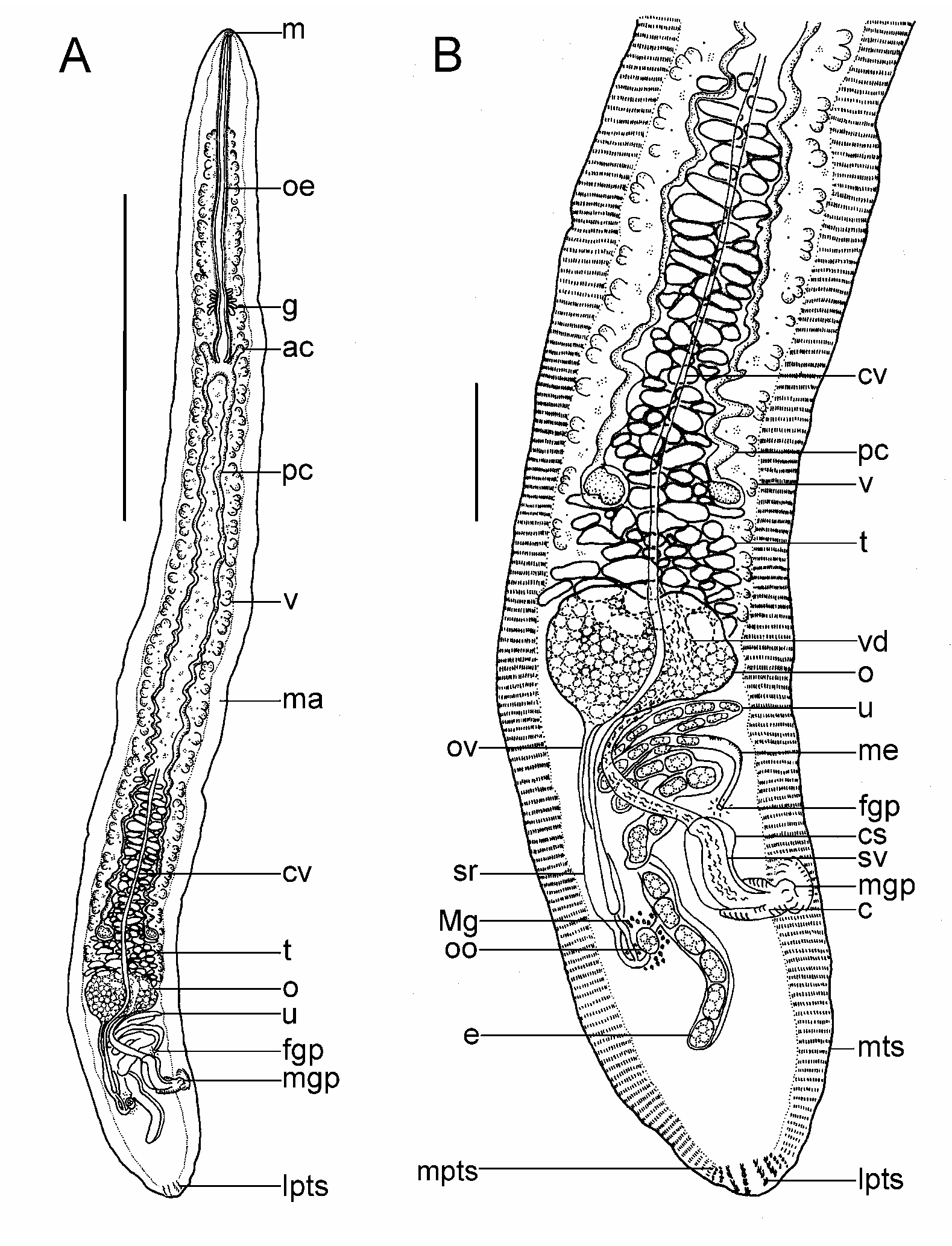
Paradeontacylix godfreyi n. sp. ( Figs 1 View FIGURE 1 , 2A View FIGURE 2 )
Typehost: Seriola lalandi Valenciennes, 1833 (Carangidae) .
Typelocality: Port Augusta, northern Spencer Gulf , South Australia (32º 42’04’’S, 137º46’17’’E) GoogleMaps .
Other locality: Killarney , Victoria (38º23’36’’S, 142º20’24’’E) GoogleMaps .
Site: Heart.
Infection details: Port Augusta: number of infected fish = 4; prevalence 12.5%; mean intensity 2 (1–5); host sizes 1410–1500 FL (350–1500 FL, n = 32). Victoria: number of infected fish = 1; prevalence 4%; intensity 1; host sizes 760 FL (460–790 FL, n = 25). The single infected specimen of S. lalandi from Victoria was also parasitised by a specimen of Paradeontacylix sp. (see below). Nodules, resulting from parasite eggs, were observed in the inner muscle layers of the heart in the ventricle and atrium.
Etymology: The species is named after Mr Reggie Godfrey, who has studied the behavioural patterns of yellowtail kingfish at Port Augusta for over 50 years. His knowledge and skill greatly assisted host and parasite collections at Port Augusta.
Material deposited: Holotype SAMA AHC 28903, 5 paratypes SAMA AHC 28904– 28908 ; 2 paratypes USNPC No. 097276.
Description
Paradeontacylix sensu Smith (2002) . Description and measurements based on 8 mature adult specimens. Body slender, dorsoventrally flattened, 4,080 (3,739 –4,215, n = 6) long by 428 (357–566, n = 8) wide; approximately 10 times longer than wide, width consistent throughout most of specimen only narrowing at anterior and posterior extremities ( Fig. 1A View FIGURE 1 ). Lateral body margins slender, skirt–like ( Fig. 1A View FIGURE 1 ), bearing numerous transverse rows of marginal tegumental spines spanning entire length of parasite except for anterior extremity. Marginal tegumental spines ventrolateral, arranged in numerous transverse rows, 816 (690–890, n = 6) (Table 1) on both sides of body, rows regularly spaced ( Fig. 1B View FIGURE 1 ); number of spines per row increasing from 4 posteriorly to 22 at mid–body region. Posteriorly, 16 large tegumental spines, 33 (22–40, n = 16) long by 4 (2– 6, n = 16) wide at base; conspicuous, claw–like distally, arranged in 4 longitudinal rows each comprising 4 spines ( Fig. 2A View FIGURE 2 ). Additionally, 2–4 rows of medium–sized spines 19 (11–23, n = 8) long by 3 (2–5, n = 8) wide at base, arranged on either side of large spines ( Fig. 2A View FIGURE 2 ).
Mouth subterminal. Oesophagus narrow anteriorly, widening posteriorly, 1,171 (928– 1,572, n = 6) long, ~30% of total body length, surrounded by gland cells along entire length, larger gland cells forming compact mass in posterior portion ( Fig. 1A View FIGURE 1 ). Short anterior and elongate posterior intestinal caeca forming H–shape; anterior caeca 74 (68– 85, n = 4) long extending anterolaterally from midline; posterior caeca often highly sinuous, 2,237 (2,050 –2,703, n = 7) long, ~56% of body length, approximately 34 times longer than anterior caeca, terminating blindly among testicular field, anterior to ovary ( Figs 1A, B View FIGURE 1 ).
Testes 99 (n = 1), mostly transversely elongate, some rounded, stacked irregularly mostly between posterior extremities of caeca. Testicular field 852 (642–1,112, n = 4) long representing 21 (18–25)% of total body length; posteriorly, overlapping anterior third of ovary ( Fig. 1A View FIGURE 1 ). Vas deferens descending at mid–line from posterior region of testicular field, passing dorsal to ovary, following curved path to form seminal vesicle filling entire cirrus sac. Cirrus cylindrical, 60 (47–75, n = 4) long. Male genital pore dorsal, near left body margin, 461 (343–569, n = 7) from posterior end of body ( Fig. 1B View FIGURE 1 ).
Ovary heartshaped, overlapped anteriorly by testes, 196 (149–279, n = 8) long and ~4% of body length, 330 (222–443, n = 8) wide or 76% of body width, located 696 (438– 959, n = 7) or ~17% of body length from posterior end of body ( Fig. 1B View FIGURE 1 ). Oviduct originating from right side of ovary, passing posteriorly, dilating to form seminal receptacle, 199 (139–261, n = 6) long, 38 (17–53, n = 7) wide. Posteriorly, seminal receptacle narrows, receives common vitelline duct, turns left to join oötype 55 (41–64, n = 7) long by 49 (36–56, n = 7) wide, ovoid, near level of cirrus sac, surrounded by Mehlis’ glands ( Fig. 1B View FIGURE 1 ). Uterus descending 137 (92–202, n = 7) posterior to oötype, then ascending through several coils filling space immediately posterior to ovary, finally descending to form slender metraterm. Female genital pore opening dorsally, anterior to male pore, at level of junction of vas deferens with seminal vesicle ( Fig. 1B View FIGURE 1 ). Distance between male and female pores 152 (84–206, n = 4). Eggs ovoid 38 (3641, n = 4) long, 30 (2934, n = 4) wide, measured in utero. Vitellarium follicular, follicles extending from level onethird along length of oesophagus posteriorly to anterior margin of ovary ( Fig. 1A View FIGURE 1 ). Common vitelline duct first observed medianly, just anterior to testicular field, passing posteriorly ventral to testes and ovary, terminating at level of oötype. Excretory vesicle and pore not observed.
Remarks
Smith (2002) provided a key to the genera of the Sanguinicolidae and revised the generic diagnosis for Paradeontacylix . According to the revision, Paradeontacylix species possess 19 to 71 testes. However, the new species described here has up to 99 testes. The number of testes in most specimens we collected was difficult to determine with accuracy, except in the holotype. While the greater number of testes is noteworthy, we have not revised the generic diagnosis here based on this single character. Testes number can be highly variable in some species, and degeneration of the testes has been observed in adult specimens of Aporocotyle simplex and Cruoricola lates (see Thulin 1980, Herbert et al. 1994). A revision may be necessary when more material of P. godfreyi becomes available. Table 1 presents a comparison of the important morphological characters of Paradeontacylix species reported from Seriola species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paradeontacylix godfreyi
| Hutson, Kate S. & Whittington, Ian D. 2006 |
Paradeontacylix
| sensu Smith 2002 |