
Malenchus geraerti, Pedram & Soleymanzadeh & Pourjam & Mobasseri, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4369.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:88E4F64A-3C03-4CC9-9937-A682AA474C81 |
|
DOI |
https://doi.org/10.5281/zenodo.5960195 |
|
persistent identifier |
https://treatment.plazi.org/id/03C9D830-1421-0F09-FF48-FCE234E9FF2C |
|
treatment provided by |
Plazi |
|
scientific name |
Malenchus geraerti |
| status |
sp. nov. |
Malenchus geraerti n. sp.
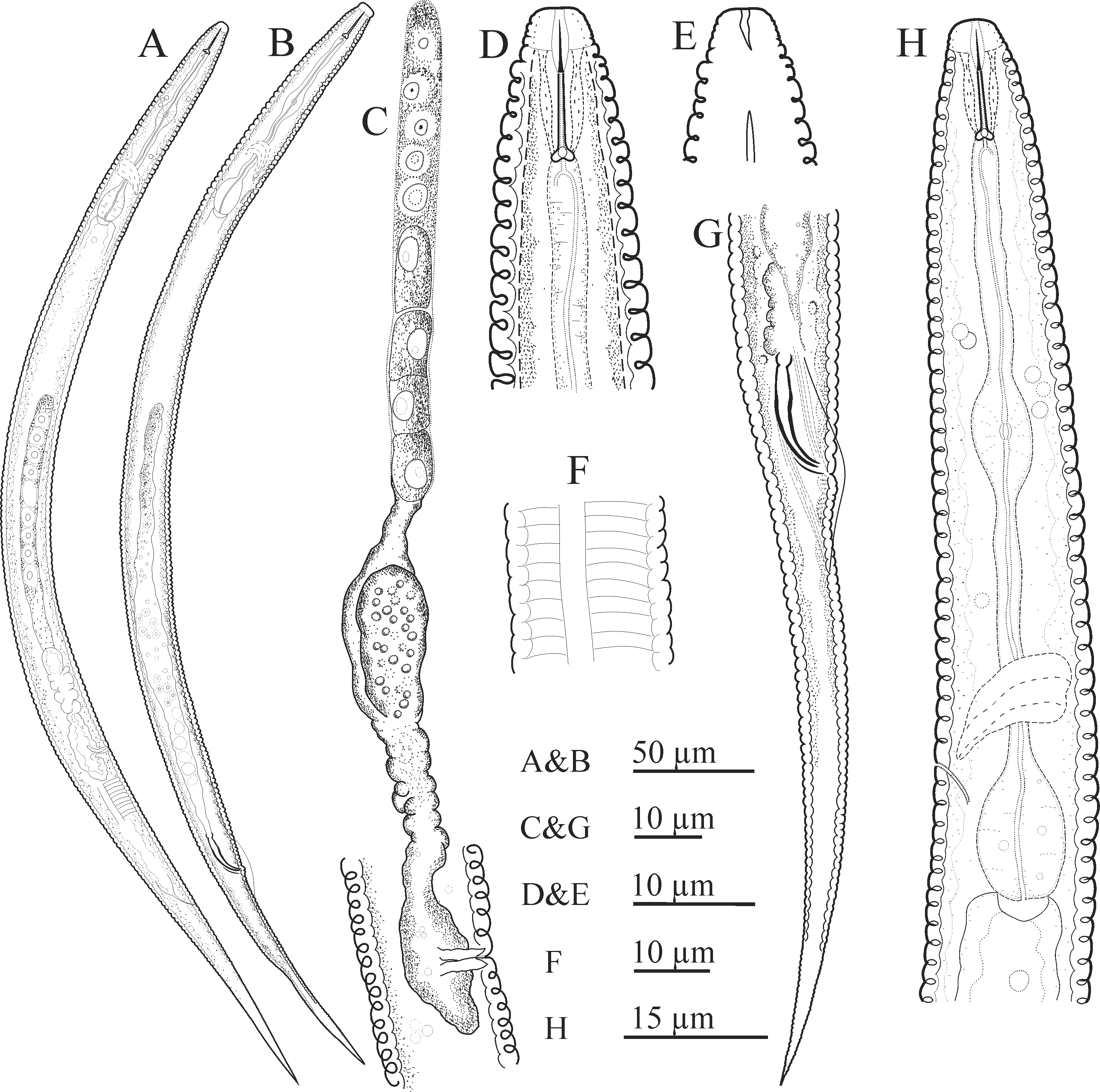
Figs 1–3 View FIGURE 1 View FIGURE2 View FIGURE 3
The new species is named after Prof. Etienne Geraert, the pioneer and outstanding scientist in the taxonomy of Tylenchidae .
Measurements, see Table 1.
Female. Body slightly ventrally curved after fixation, markedly narrowing from the level of vulva to distal end. Cuticle coarsely annulated, annuli 2.1–2.3 µm wide at mid-body. Lateral field a plain band ( i.e. composed of two incisures), raised in cross section, constricted at the junction with body ( Fig. 3B View FIGURE 3 , arrows), originating at about the level of the middle of the stylet shaft in its resting position. Lip region continuous with body contour; not conspicuously narrower than the adjacent body, 5–7 µm wide, 3–4 µm high, bearing 3–4 annuli. Amphidial openings S-shaped slits, starting from labial plate, extending posteriorly, markedly narrowing and reaching base of the head (extending to the weakly sclerotized cephalic framework). Stylet shaft moderately developed, conus much finer, knobs directed posteriorly. Dorsal gland orifice (DGO) close to knobs. Excretory pore (ex. pore) at the level of pharyngeal bulb, slightly variable in position, 91–94 µm from anterior end. Hemizonid 92 µm ( n = 3) and nerve ring 63–75 µm from anterior end. Pharynx with slender procorpus, muscular, rounded to ellipsoid metacorpus with well-developed valve and corresponding plates at 43–49 µm from anterior end, slender/narrow isthmus and saccate pharyngeal bulb 13–18 × 8–11 µm. Reproductive system mono-prodelphic, ovary outstretched, oocytes mostly in single file, except in germinal zone, oviduct short, spermatheca elongate-rounded, filled with small spheroid sperm cells, offset and sometimes appearing as bilobed (with depression or constriction at middle), crustaformeria and uterus not clearly discernible from each other. Vagina perpendicular to body axis, with moderately sclerotized walls. Small, undifferentiated postvulval uterine sac (PUS) present. Vulva sunken inside body, in the form of a transverse slit, with large epiptygma and no flaps. Prophasmid barely visible, located 14 µm anterior to vulval opening (n = 1). Vulva–anus distance 64–77 µm, or 0.8–1.1 times the tail length. Tail conical, tapering gradually to a more or less pointed tip.
Male. About half as frequent as females in the studied population. General morphology similar to that of female, except in sexual characters. Testis single, outstretched. Spicules tylenchoid, narrow and slender, slightly arcuate distally, 17–22 µm long with rounded capitulum, slender shaft and blade narrowing toward distal end. Gubernaculum small, 4–6 µm. Bursa cloacal, 26–43 µm long, with smooth margin. Tail similar to that of female.
Type Habitat And Locality. Recovered from soil samples collected about rhizosphere of forest trees in forests of Gilan province, northern Iran. GPS coordinates: 37°7ʹ30.719ʹʹN, 49°39ʹ25.742ʹʹE.
Type Material. Five females ( holotype and paratypes) and five males deposited in the Nematode Collection of the Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran. Five paratype females and two males deposited at each of the following collections: WANECO collection, Wageningen, The Netherlands, and UGent Nematode Collection of the Nematology Research Unit, Department of Biology, Ghent University, Ghent, Belgium.
Diagnosis And Relationships. The new species belongs to the subgenus Malenchus owing to its amphidial opening characteristics (S-shaped in lateral view, wide at anterior end, narrowing distally and reaching to the base of the head), and two lateral lines, forming a plain band under light microscopy. It is further characterized by having short-bodied females, lateral field origin at about the middle of the stylet shaft, muscular, rounded to ellipsoid metacorpus with well-developed valve and valvular plates, small PUS, vulva sunken in body with large epiptygma and no flap, conical tail, tapering gradually to a more or less pointed tip, functional males with 17–22 µm tylenchoid spicules, adcloacal bursa with smooth margin and tail similar to that of female. Morphologically, the new species is similar to five known Malenchus species: M. fusiformis ( Thorne & Malek, 1968) Siddiqi, 1979 , M. machadoi ( Andrássy, 1963) Andrássy, 1968 , M. pachycephalus Andrássy, 1981 , M. solovjovae Brzeski, 1989 and M. undulatus but most closely resembles M. pachycephalus forming a morphologically-similar cryptic species pair.
Compared to M. fusiformis , a poorly known species of the genus described based only on one female and only known from its type locality ( Geraert 2008), the new species has a longer body (439–489 vs 350 µm), narrower annuli (2.1–2.3 µm wide at mid-body vs 3 µm), posteriorly directed stylet knobs ( vs rounded) and lacks vulval membranes ( vs having obscure lateral membranes).
Compared to M. machadoi , the new species has a shorter body (439–489 vs 520–560 µm), lip region continuous with body contour ( vs markedly narrower), annuli 2.1–2.3 µm wide at mid-body ( vs 1.6–1.8 µm) and median bulb with distinct valves ( vs very small or indistinct valves, as described by Andrássy 1968).
Compared to M. pachycephalus , besides differences in partial SSU and LSU rDNA sequences and divergent placement in the corresponding phylogenetic trees, the new species differs in the nature of the median bulb (welldeveloped with valvular plates vs very weakly developed, without valve), stylet knobs (symmetrical vs slightly asymmetrical), lateral field margin (smooth vs crenate), pharynx length (93–108 vs 86–90 µm) and lacks vulval flaps ( vs small dikes present). Furthermore, the raised lateral fields of the new species are separated from the rest of the body by a sharp constriction ( Fig. 3B View FIGURE 3 , arrows); this demarcation is lacking in M. pachycephalus as illustrated by Qing et al. (2017).
Compared to M. solovjovae , the new species has a well-developed stylet and corresponding knobs and welldeveloped valve in the median bulb.
Compared to M. undulatus , besides differences in SSU and LSU rDNA sequences and distant positions in the corresponding phylogenetic trees, the new species has annuli 2.1–2.3 µm wide ( vs 1.7–1.9), plain lateral field margins ( vs deeply crenate), lip region continuous with body contour ( vs distinctly narrower), longer stylet (11–13 vs 9.5–10.0 µm), more posterior vulva (V = 66.0–69.5 vs 63–66) and lacks vulval dikes ( vs having dikes 3 annuli long).
Compared to M. neosulcus Geraert & Raski, 1986 , its sister taxon in our SSU rDNA phylogeny, the new species has 3–4 head annuli ( vs 5–6), lateral field with plain margins ( vs slightly crenate), well-developed median bulb, valve, and valve plates ( vs weakly developed), wider body annuli (2.1–2.3 vs 1.8–2.0 µm), shorter tail (65–87 vs 86–96 µm), with smaller c' (5.9–8.1 vs 8–10) and greater c (5.4–6.8 vs 4.7–5.5) values, and differences in SSU sequence (accession numbers KX907138 View Materials for the new species and KJ869330 View Materials for M. neosulcus ).
Compared to M. labiatus , its sister taxon in our LSU rDNA phylogeny, the new species has a longer stylet (11– 13 vs 8–9 µm), greater V value (66.0–69.5 vs 59–63), shorter tail (65–87 vs 98–112 µm) and lacks both vulval flaps and a disc-like structure at the lip apex.
Molecular Phylogeny. Partial sequencing of the SSU rDNA gene from the new species yielded a single fragment 1066 nt long. Blast searches using the sequence of this fragment revealed the highest coverage with two sequences of M. neosulcus ( KJ869330 View Materials ) and M. acarayensis Andrássy, 1968 ( KJ869331 View Materials ), with identities of 93% and 91%, respectively. A 98% identity was achieved for a short (74% coverage) sequence of M. pachycephalus . Two longer sequences (with 88% coverage) of M. pachycephalus ( KX156286 View Materials and KX156286 View Materials ) had 92% identity with partial SSU rDNA of the new species. Lower identity and coverage percentages were returned for other sequences. A total of 98 species/isolates of Tylenchoidea Ӧrley, 1880, along with Aphelenchoididae Skarbilovich, 1947 and two panagrolaimid species as outgroup taxa were selected for our SSU rDNA-based phylogenetic analysis. This dataset comprises 1495 total characters, of which 739 are variable. Figure 4 View FIGURE 4 presents the Bayesian phylogenetic tree inferred using this dataset. The major clade of Malenchus spp. in this tree (based on currently available SSU sequences for the genus, and designated M, in green) has full BPP and moderate (62%) ML BS support. The new species forms a well-supported clade (1.00 BPP / 98% MLBS) with M. neosulcus in this tree. The phylogenetic relationship of these two species ( M. geraerti n. sp. and M. neosulcus ) with M. acarayensis , M. pachycephalus and an unidentified isolate of Malenchus sp. ( KX156289 View Materials ) is not further resolved due to polytomy.
Partial sequencing of the LSU rDNA gene (D2/D3 region) from two females of the new species yielded single fragments of 650 (isolate1) and 654 (isolate 2) nt long. A Blast search using these fragments’ sequences indicated no significant identity with any of the currently available sequences deposited in GenBank. A total of 90 species/ isolates of Tylenchoidea (ingroup members) and Aphelenchoididae (as outgroup taxa) were selected for our LSU rDNA gene (D2/D3 region) phylogenetic analysis. This dataset contains 468 total characters, of which 297 are variable. Figure 5 View FIGURE 5 presents the Bayesian tree inferred using this dataset. Species of the genus Malenchus occur in two major clades, A and B, in this tree. Clade A also includes the ambiguous species Filenchus balcarceanus Torres & Geraert, 1996 ( sensu Qing et al. 2017), which shares characters of both Filenchus Andrássy, 1954 and Malenchus . The monophyly of this clade is highly supported (1.00 BPP / 81% MLBS). Clade B is a strongly supported (1.00 BPP / 99% MLBS) monophyletic group including our new, three known, and three unidentified isolates/species of Malenchus , along with two species of Lelenchus Andrássy, 1954 . The new species clusters in a subclade containing two species of Lelenchus ( KU234169 View Materials , KP730042 View Materials , KX156335 View Materials ) and M. labiatus ( KP730047 View Materials ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |