
Pachycerianthus delwynae, Carter, 1995
|
publication ID |
https://doi.org/10.3853/j.0067-1975.47.1995.3 |
|
DOI |
https://doi.org/10.5281/zenodo.4659350 |
|
persistent identifier |
https://treatment.plazi.org/id/03C987E8-FFE9-FF93-17D0-7A5BFA54ADC9 |
|
treatment provided by |
Felipe |
|
scientific name |
Pachycerianthus delwynae |
| status |
sp. nov. |
Pachycerianthus delwynae n.sp.
Type material. HOLOTYPE: Port Jackson , Chowder Bay, muddy sand, 14 m, coIl. S. Carter, 15 September 1989, AMG15399 . PARATYPES ( 2 specimens): Port Jackson , Bottle & Glass Rocks, sandy mud, 5 m, colI. P. Watts, 13 July 1989, AMG15400 ; Bottle & Glass Rocks, amongst mussel shell debris, 7 m, colI. S. Carter, 20 July 1989, AMG15401 . Additional material ( 1 specimen): Port Jackson , Manly pool, colI. Dr P. Groves, 31 December 1957, AMG15404 .
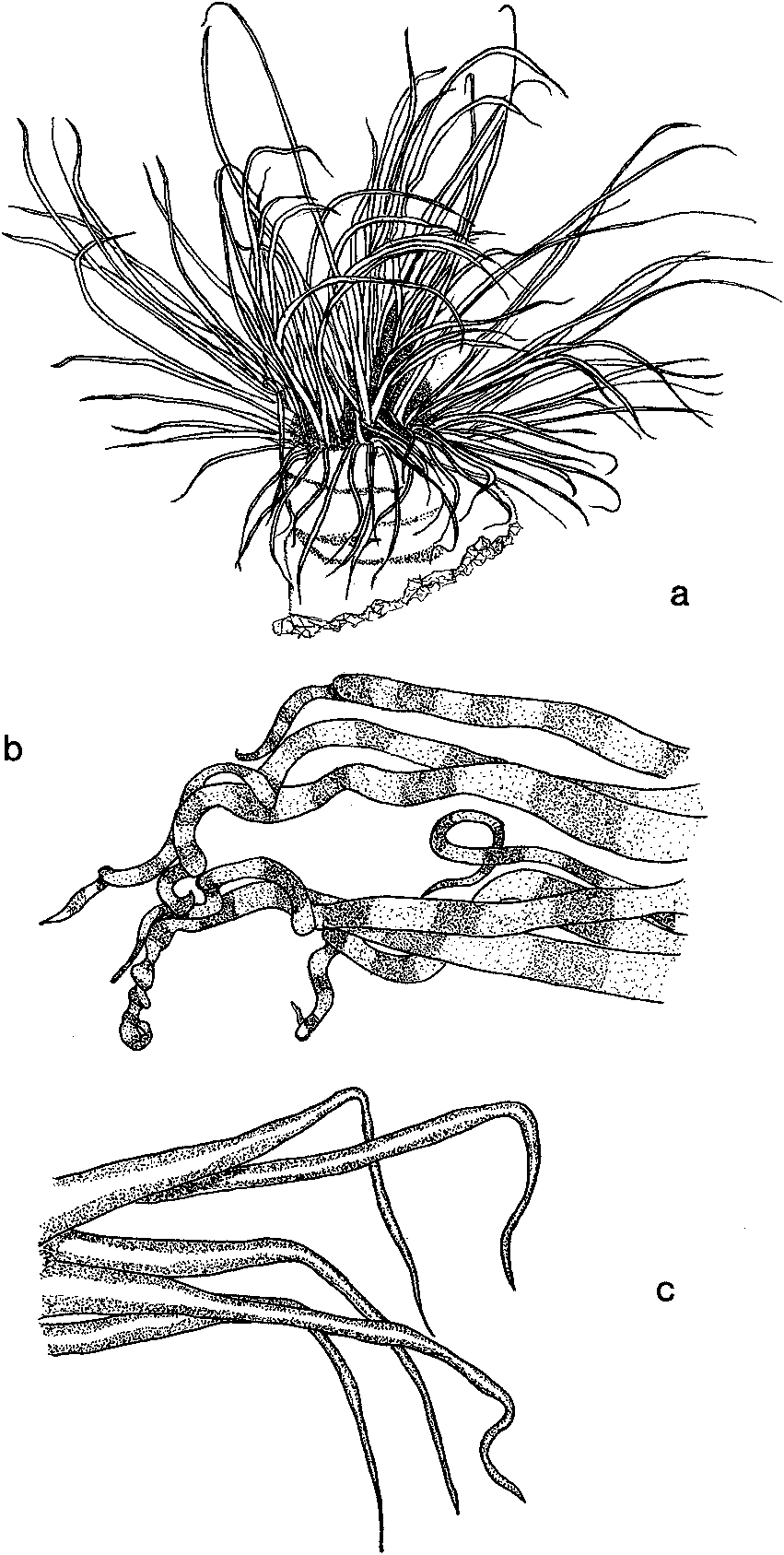
Description. Coloration of live animals. Column purplish brown aborally, merging through dark to pale yellowish brown to dark brown just below oral disc. Marginal tentacles pale orange to reddish brown with indistinct pale or white bands. Labial tentacles pale yellowish brown to cream colour. The oral disc is purplish brown.
Coloration ofpreserved animals. Purple colour faded leaving overall brown coloration. Marginal tentacles still possess distinct pale bands. Labials are pale yellowish brown.
Length in preservative. 50-240 mm.
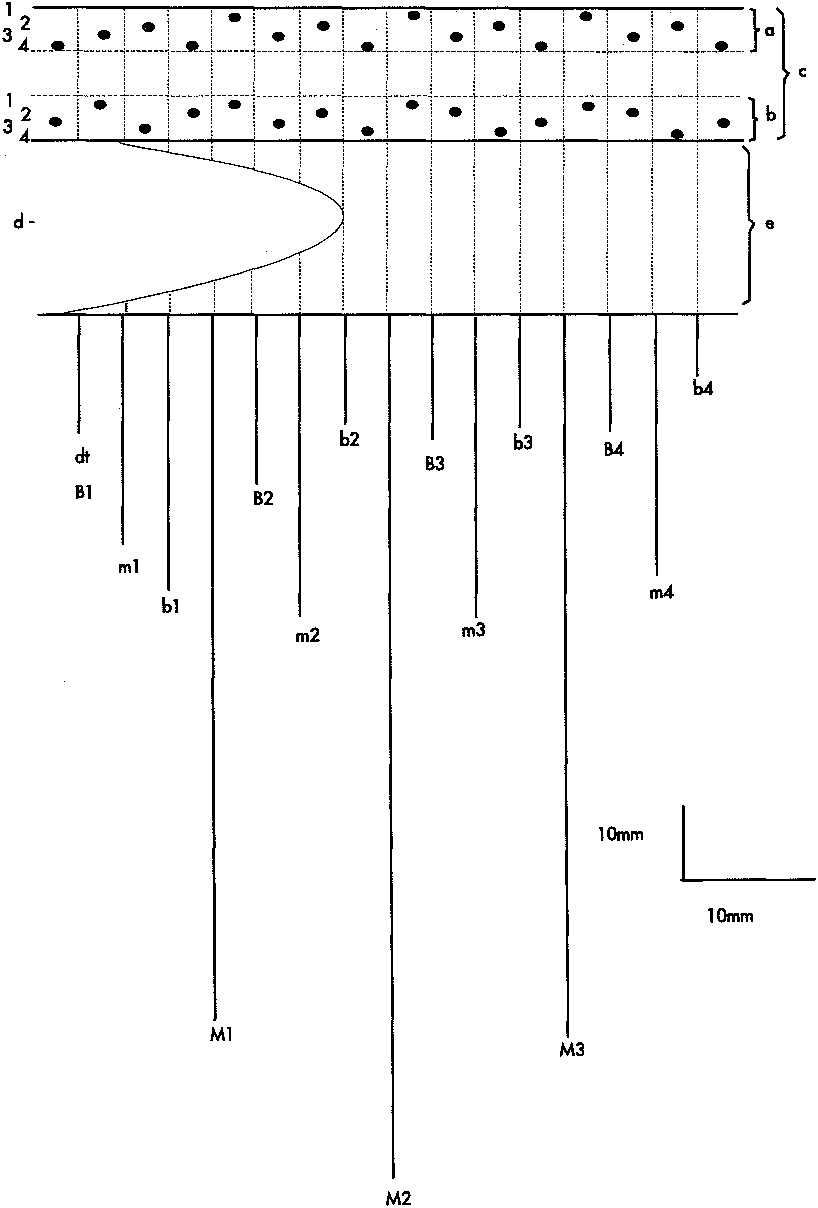
Tentacles. Formula for arrangement of tentacles into pseudocycles are: marginal tentacles, 1(dt)2314. 2314.2314... labial tentacles, 2(dt)413.4231. 4312.4312... or 1(dt)341.4231.4312... In each crown, division into pseudo-cycles is difficult in some specimens. Directive labial tentacle (dt) is present. Marginal tentacles number 89-114 (type 110). Labial tentacles number 89-112. When animal not actively feeding, but still extended from tube, distal half to one third of marginal tentacles hang limply with distal quarter coiled loosely. This distal quarter remains coiled when animal withdraws into its tube.
Siphonoglyph. Running two-thirds length of I\ctinopharynx. Attached mesenteries 6 or 7 ( 7 in type). Hyposu1cus well developed, reaching one fifth length of siphonoglyph. Hemisu1cus distinct, continuing down directive tentacle.
Mesenteries. Directive mesenteries (B1) shorter than siphonoglyph; lengths of mesenteries may differ in same animal. Second mesenteries (m1) sterile, more than twice length of directives. Ciliated tract runs whole length of m1 and a few craspedonemes occupy aboral end. Third mesentery (b1) sterile, 1-1.5 x length of ml. Cnidoglandular tract found on distal half, a few craspedonemes may be present on distal end of b1 mesentery.
Mesenterial arrangement is BmbM. All macromesenteries (M, m) fertile, all micromesenteries (B, b) infertile. M1 long, but never reaches aboral pole. Three quarters to all of M1 occupied by ciliated tract and distal three quarters possess craspedonemes. The m1 mesenteries less than half length of M1 with ciliated tract running length of mesentery, distal half with craspedonemes. M2 longer or shorter than M1 and may reach aboral pole. Ciliated tract occupies nearly entire length of M2. After second quartette, M mesenteries reduced in length toward reproductive chamber, m mesenteries all of similar length. Micromesenteries usually very short. B1 usually similar in length to directive mesentery with b1 being half the length of Bl. All the micromesenteries carry considerable amounts of cnidoglandular tract. After second quartette, micromesenteries vary little in length ( Fig. 2 View Fig ).
Cnidom. Spirocysts, atrichs, homotrichs and spirulae of several types are present. The measurements were taken from 20 cnida for each type from three samples (n = 60) from two paratype specimens. All cnida are considered numerous and common unless specified.
Spirocysts appear to be absent from the column and the actinopharynx. The classes of spirulae could be separated on the length of the shaft in relation to the total capsule length. Spirulae 2 is the most common type, found in all tissue except the single chord of filament. Atrichs were found in all organs except the single chord and were uncommon in the labial and marginal tentacle tips. Homotrichs appear to be confined to the column.
Biology. Specimens were all collected from, or observed in, sand and/or muddy sediments, often with overlying banks of mussel shell (Mytilus sp) detritus or among parchment worm (Diopatra dentata) colonies. Depth range varied from three to over 15 m. The tubes rarely protruded more than three centimeters above the substrate. Several specimens have been collected with large numbers of the commensal worm, Phoronis australis , living in the tube walls. Coleman (1987) reported the presence of a commensal shrimp Periclemenes holthuisi and crab Lissocarcinus sp in association with an unidentified cerianthid which appears to be similar to Pachycerianthus delwynae .
Specimens viewed in an aquarium or in their natural habitat were rarely active unless light levels were low, suggesting that P. delwynae is crepuscular.
Remarks. Pachycerianthus delwynae possesses a directive labial tentacle, M1 never reaches the aboral pole and M2 may be longer than Ml. M mesenteries are much longer than m mesenteries. The distinctive banded marginal tentacles are commonly coiled when relaxed ( Fig. 1b View Fig ). These features distinguish P. delwynae from the other known species of Pachycerianthus .
Etymology. Pachycerianthus delwynae is named after Ms Delwyn Hunt for her support and assistance during this study.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Ceriantipatharia |
|
Order |
|
|
SubOrder |
Spirularia |
|
Family |
|
|
Genus |