
Thalassodes antithetica Herbulot, 1962
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4139.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:0BE46816-899A-4A97-878A-AC1845FC7C97 |
|
DOI |
https://doi.org/10.5281/zenodo.6072159 |
|
persistent identifier |
https://treatment.plazi.org/id/03C8F63B-FFDC-FFDA-FF7F-CEDEFF719D81 |
|
treatment provided by |
Plazi |
|
scientific name |
Thalassodes antithetica Herbulot, 1962 |
| status |
|
Thalassodes antithetica Herbulot, 1962 View in CoL
( Figs 1–14 View FIGURES 1 – 14 )
Thalassodes antithetica Herbulot, 1962: 31 View in CoL .
Thalassodes antithetica Herbulot, 1964 View in CoL [erroneous year, the description was published in 1962]: Legrand 1966: 129; Lawrence 2005: 95; Gerlach and Matyot 2006: 67.
Material examined. Holotype ( MNHN), ♂, Seychelles: Mahé, B. Vallon, 14.vi.1960, H. Legrand (Pr. №4269, C. Herbulot). Paratype (designated as allotype by C. Herbulot) ( MNHN), ♀, Seychelles: Mahé, Misere, v.1960, A. Cauvin (138). Seychelles: Praslin, Anse Possession, 418ʹ36ʺ S, 5543ʹ45ʺ E, 5 m alt., coastal lowland, 5–8.i.2013, I. Bolotov leg., 1♀ ( RMBH: voucher no. INEP-g709); Praslin, Anse Boudin, main road, junction to Zimbabwe, 417'55''S, 5542'35''E, 25 m alt., lowland forest, 5–9.vii.2014, M. Bippus leg., 1♀, 6♂ ( PCMB); Mahé, Au Cap, 442ʹ59ʺ S, 5531ʹ14ʺE, 40 m alt., disturbed mid-altitude forest dominated by palms and cinnamon trees on a hill slope, 24.i.2013, I. Bolotov leg., 1♀ ( RMBH: voucher no. INEP-g710).
Diagnosis. This species can be distinguished from all congeners by long and narrowed valva with a costal extension to form a sclerotized curved free arm, as well as the presence of two long lateral processes on the posterior margin of the male eighth sternite. Based on the male genitalia structure and smooth octavals, the species is similar to Pelagodes bellula Han & Xue, 2011 , a species of another genus, but differs by much more narrow and elongated costal extension of the valva. Additionally, the species differs from all Pelagodes taxa by a prominent white discal spot on the hind wing and notable creamy white costal margin of the upper wing.
Redescription. Head: Male antenna bipectinate with filiform terminal part; female antenna filiform. Frons bluish green. Labial palpi short, barely extended beyond the frons, yellowish white. Vertex and stem of antennae pure white. Thorax: Dorsal side of thorax deep bluish green, ventral side whitish. Legs yellowish white, inside of foreleg reddish. Hind tibia has a pair of spurs. Forewing length: ♂ 14.5–16.5 mm (14.5 mm in holotype); ♀ 14.3–17.0 mm (15.5 mm in paratype). Venation is typical for Pelagodes , Thalassodes and other closely related genera ( Holloway 1996; Han & Xue 2011, 2011a): Discal cell shorter than 1/2 length of wings. Forewing with R1 diverging from upper angle of cell, connected with Sc; R2–5 shortly stalked with M1, M3 shortly stalked with CuA1. Hindwing has Rs stalked with M1, M3 stalked with CuA1; discocellulars are oblique; 3A is absent. Upperside of wings uniformly deep bluish green; rarely diffused with unclear white striations; with creamy white costal margins to both wings; hind wing with a prominent white discal spot. Herbulot (1962) did not mentioned this spot in his protologue, possibly because of the relatively poor condition of the type specimens. Underside of wings pale beige. Fringes of both wings yellowish white. Angle on outer margin of the hind wing relatively weakly sharpened, rounded. Abdomen: Upperside bluish green, underside whitish. In male, the eighth tergite slightly concave at the posterior margin; the eighth sternite with two very long, smooth, curved sclerotized lateral processes (octavals). Male genitalia: Uncus rod-like, tapering posteriorly. Socii relatively narrow and tapering. Gnathos with sclerotized simple median process. Valva long with broad basal half and narrow distal half; costal extension slender, smooth, acute, not projecting beyond the apex of valva. Juxta with two sclerotized lateral processes on posterior margin. Saccus semicircular. Phallus (aedeagus) short, thick, expanded at the middle, phallus surface sclerotized posteriorly. Coremata absent. Female genitalia: Ductus bursae short, very broad and sclerotized; the corpus bursae rounded or oval, three times longer than ductus bursae. Signum absent.
DNA barcoding. Molecular analysis was performed as described in Bolotov et al. (2014). We obtained two sequences of the cytochrome oxidase subunit I (COI) gene, resulting in an 816-bp fragment (NCBI’s GenBank acc. nos. KM 111177 View Materials and KM 111178 View Materials for specimens from Praslin and Mahé, respectively). Both sequences belong to a single haplotype, providing support for the hypothesis that there has been recent natural or human-mediated movement of Lepidoptera across the approximately 45 km stretch of sea between Praslin and Mahé ( Bolotov et al. 2015). We were unable to find the COI sequences of closely related taxa in the NCBI’s GenBank using a Basic Local Alignment Search Tool (BLAST). Thus, phylogenetic relations of the haplotype was retrieved with the BOLD systems COI Full Database, which includes records without species designation. The most similar sequence belongs to Thalassodes sp. AH01Au (sample ID: BC ZSM Lep 10205), an emerald moth specimen from Queensland, Australia, with p -distance 3.1% (Appendix 1). Among the other top 10 nearest neighbor sequences (p -distance ranges from 3.9 to 4.6%) moth specimens from Indonesia, Philippines and Thailand are also presented. Additionally, the top 99 matches from BOLD database belong almost exclusively to specimens from the Indo-Australian tropics, excluding four Thalassodes specimens from Africa, which share the most distant haplotypes (Appendix 1). This preliminary assessment reveals that the Seychelles species is closely linked to the Indo-Australian fauna. DNA barcoding of Sauris mouliniei (Legrand, 1971) , another Seychelles endemic geometrid species, also reveals its closest affinity to be with Australian congeners ( Bolotov et al. 2014). However, this was only a preliminary report because the phylogenetic relationships of this species with supposedly related African taxa (e.g. genus Episteira and others) are unknown.
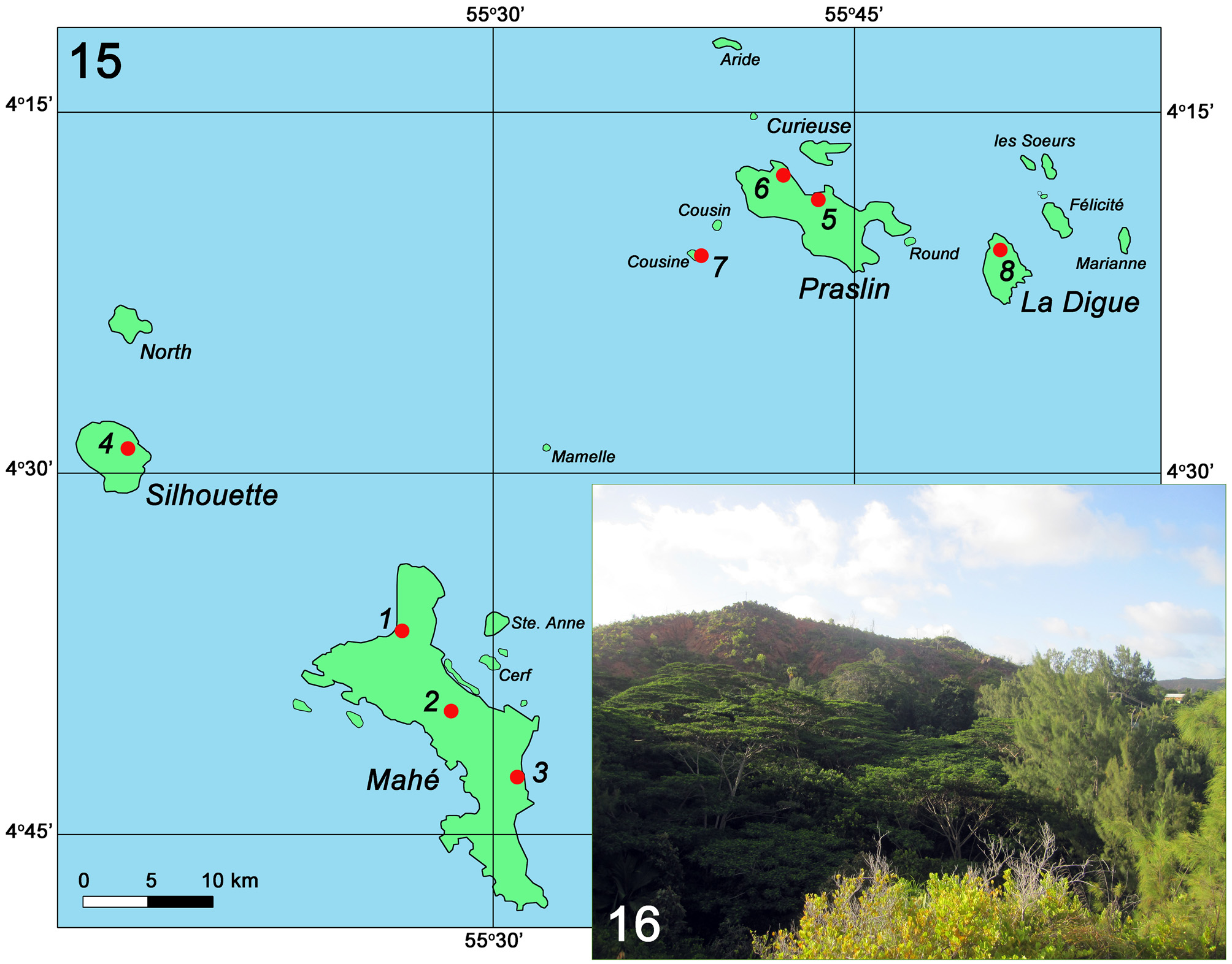
Distribution and ecology. The species is considered to be endemic of the granitic Seychelles ( Fig. 15 View FIGURES 15 – 16 ) and examples have been collected from Mahé, Silhouette ( Herbulot 1962; Legrand 1966; our data), Cousine ( Lawrence 2005), La Digue ( Mazzei 2014: documented by photos of five live specimens) and Praslin (our data: first record for the island). The species inhabits coastal lowlands and mid-altitude forests. The species was very rare in the majority of collection localities, excluding forest sites at Anse Boudin, Praslin ( Fig. 16 View FIGURES 15 – 16 ) and near the Calou Guest House, La Digue, where it was recorded as relatively common (M. Bippus and P. Mazzei, pers. comm.). It has been observed by us resting during the day on the underside of leaves of Chrysobalanus icaco (L.) L. near Beau Vallon on Mahé, the green colour of the wings clearly having cryptic value against the green background of the leaves (P. Matyot, pers. comm.). Adult specimens were found almost all year round ( Herbulot 1962; Lawrence 2005; Mazzei 2014; P. Mazzei, pers. comm.; our data).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Thalassodes antithetica Herbulot, 1962
| Bolotov, Ivan N., Matyot, Pat, Bippus, Maik, Spitsyn, Vitaly M., Kolosova, Yulia S. & Kondakov, Alexander V. 2016 |
Thalassodes antithetica
| Gerlach 2006: 67 |
| Lawrence 2005: 95 |
| Legrand 1966: 129 |
Thalassodes antithetica
| Herbulot 1962: 31 |