
Manayunkia occidentalis, Atkinson & Bartholomew & Rouse, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4751.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:7B8087E5-4F25-45F2-A486-4521943EAB7E |
|
DOI |
https://doi.org/10.5281/zenodo.3718189 |
|
persistent identifier |
https://treatment.plazi.org/id/62A9E61E-4428-4EA5-9BAA-9D9621B5F481 |
|
taxon LSID |
lsid:zoobank.org:act:62A9E61E-4428-4EA5-9BAA-9D9621B5F481 |
|
treatment provided by |
Plazi |
|
scientific name |
Manayunkia occidentalis |
| status |
sp. nov. |
Manayunkia occidentalis View in CoL sp. nov.
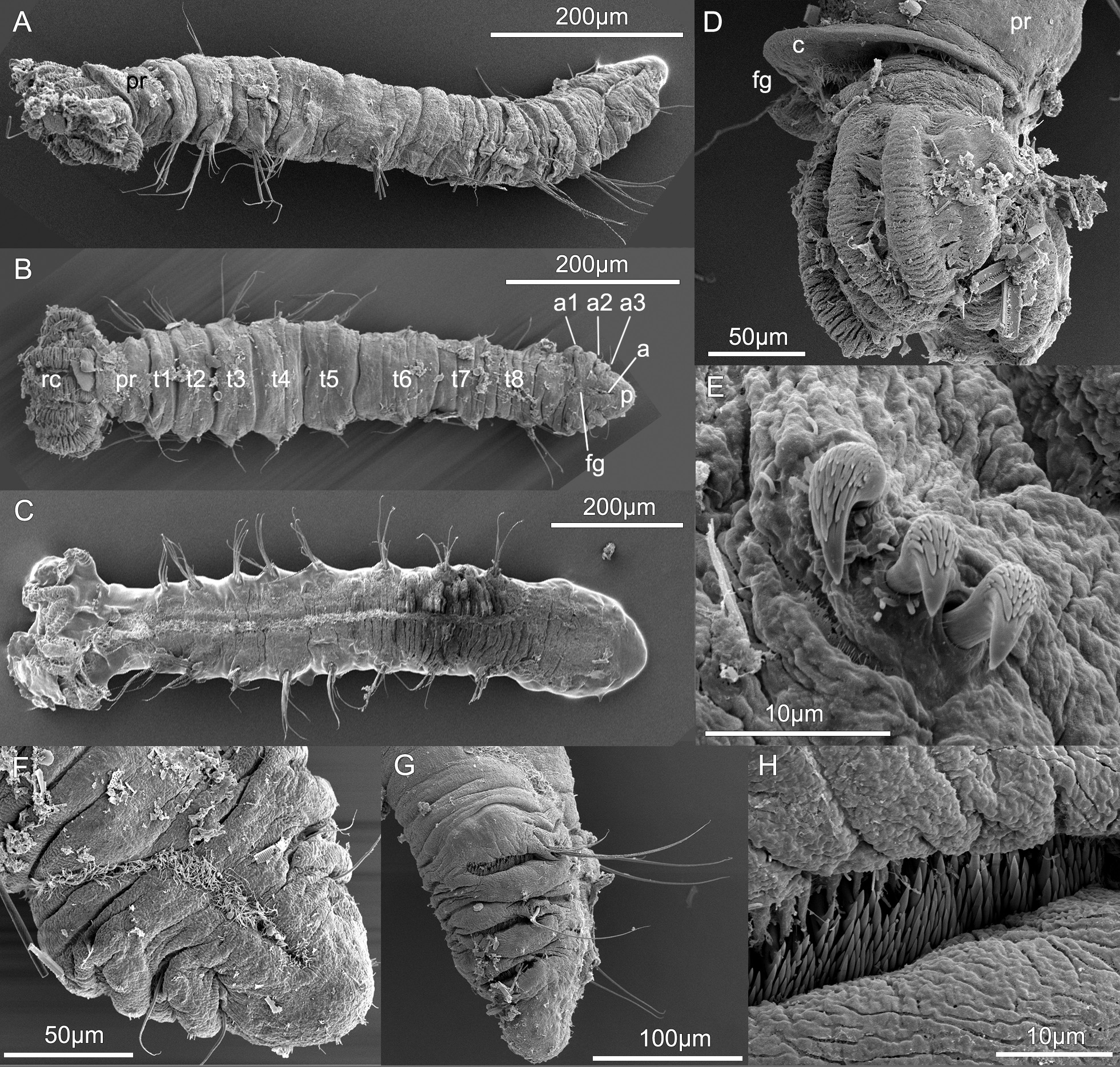
Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4
Manayunkia speciosa: Hazel, 1966 View in CoL ; Bartholomew et al. 1997; 2006; 2007; Alexander et al. 2014; Stocking and Bartholomew, 2007; Bjork & Bartholomew 2009; Willson et al. 2010; Atkinson et al., 2011; Malakauskas et al., 2013; 2015.
Holotype (in 70% ethanol, initially fixed in 10% formalin). Adult male from Klamath River mainstem near confluence with Beaver Creek, California, USA. 41.867240°N, - 122.809763°E, collected by Julie Alexander ( OSU), 25 April 2014 ( SIO-BIC A12115 View Materials ) GoogleMaps
Paratypes (in 70% ethanol, fixed in 10% formalin); One specimen, adult female, same locality, collector and date as holotype ( SIO A12116 View Materials ); One specimen, adult male, same locality, collector and date as holotype ( SIO A12117 View Materials ) .
Etymology. The specific epithet is derived from the Latin word for “west”, to contrast with M. speciosa , which occurs in eastern and central North American drainages.
Description. Holotype (SIO-BIC A12115 View Materials ) a male with eight thoracic segments and three abdominal segments. Total length (unfixed) 3.0 mm, crown 0.3 mm. Body cylindrical with tapering, dorso-ventrally flattened abdomen. Body wall translucent grey with minor brown pigmentation on peristomium and pygidium, and green pigmentation on chaetiger 6. Branchial crown ~10% of total body length. Two pairs of radioles, each with six pinnules. One ventral pinnule on radiole with 4–5 yellow-white spots in life ( Fig. 2 View FIGURE 2 B–C). One pair of vascularised, unbranched ventral filamentous appendages present, extending for about two-thirds length of radiolar crown, approximately same width as pinnules ( Fig. 2 View FIGURE 2 B–C). Anterior peristomial ring with membranous collar, followed by posterior peristomial ring ( Fig. 4 View FIGURE 4 A–B,D). Collar margin smooth, higher ventrally, separated by a narrow dorsal gap that corresponds with ciliated faecal groove ( Fig. 4D View FIGURE 4 ). Peristomial eyes, black rounded spots ( Fig. 2E View FIGURE 2 ). Pygidial eyes absent. Superior thoracic notochaetae elongate, narrowly hooded; 5–7 per fascicle (chaetigers 1–5), 4–5 per fascicle (chaetigers 6–8) ( Figs 3A View FIGURE 3 ; 4 View FIGURE 4 A–C). Inferior thoracic notochaetae on chaetigers 1–8 short; 3–4 per fascicle except chaetiger 2, which has 6 per fascicle. Thoracic neuropodia each with 4–7 uncini in chaetigers 2–8 ( Fig. 3B View FIGURE 3 ). Thoracic uncini with rows of evenly small teeth gradually decreasing in size away from main fang ( Figs 3B View FIGURE 3 ; 4E View FIGURE 4 ). Position of chaetae and uncini reversed in abdominal segments; number of abdominal uncini per fascicle decreasing posteriorly from 22 to 9. Abdominal uncini each with multiple rows of teeth that are uniform in size ( Figs 3C View FIGURE 3 ; 4H View FIGURE 4 ); manubrium at least five times longer than dentate region, with base about two thirds the width of the dentate region. Abdominal neuropodia are elongate, narrowly hooded, decreasing posteriorly from 4–6 to 2 per fascicle ( Fig. 4G View FIGURE 4 ).
Variation. Paratypes are similar to the holotype. However, females have oocytes in thoracic chaetigers 4–5 ( Fig. 2C View FIGURE 2 ) and a pair of pigmented spermathecae in the base of the radiolar crown. Females differ from males in having thoracic neuropodia with uncini in chaetigers 2–5 followed by 3–6 elongate hooded chaetae in neuropodia of chaetigers 6–8 ( Fig. 3E,G View FIGURE 3 ).
Remarks. Manayunkia occidentalis sp. nov. is the sister group to M. speciosa based on COI sequence data with a minimum pairwise distance (simple distance) to our M. speciosa samples of 11.0%, clearly supporting that they can be regarded as a separate taxon, although phylogenetic support for this relationship was weak (bootstrap 28%). Manayunkia occidentalis sp. nov. and M. speciosa are the only species of Manayunkia , and in fact of any Fabriciidae (see Huang et al. 2011), to show sexual dimorphism with regards to thoracic chaetal complement. Males of M. occidentalis sp. nov., and M. speciosa (Rouse pers. obs.) on specimens from near the type locality (SIO-BIC A12106 View Materials ), show the typical arrangement of thoracic uncini in each of chaetigers 2–8, while females have thoracic uncini in chaetigers 2–5 only ( Fig. 3H View FIGURE 3 ). The neuropodia of chaetigers 6–8 instead have elongate hooded chaetae ( Fig. 3G View FIGURE 3 ). These elongate hooded chaetae were seen in all females examined. Larval development would be worth studying to see if there is a loss of thoracic uncini or if they never develop in chaetigers 6–8. It is somewhat surprising that this major difference in chaetal complement was never noted previous in observations on M. speciosa . Manayunkia occidentalis sp. nov. differs markedly from descriptions of M. speciosa by Leidy (1883) and Pettibone (1953) in that it has far fewer pinnules on the radioles. In this regard Manayunkia occidentalis sp. nov. is similar to other Manayunkia species that have relatively few pinnules (see Rouse 1996).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Manayunkia occidentalis
| Atkinson, Stephen D., Bartholomew, Jerri L. & Rouse, Greg W. 2020 |
Manayunkia speciosa
| : Hazel 1966 |