
Tropidophorus laotus, Smith, 1935
|
publication ID |
https://doi.org/ 10.1206/360.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03C75B19-0E1D-1566-FD74-EC2A3A96FEC2 |
|
treatment provided by |
Tatiana |
|
scientific name |
Tropidophorus laotus |
| status |
|
laotus , Scincella melanosticta , Tropidophorus microlepis , and Cyclemys oldhamii .
These patterns of faunal overlap and divergence among northern Indochina’s upland and highland herpetofauna might be explained by three major nonexclusive hypotheses.
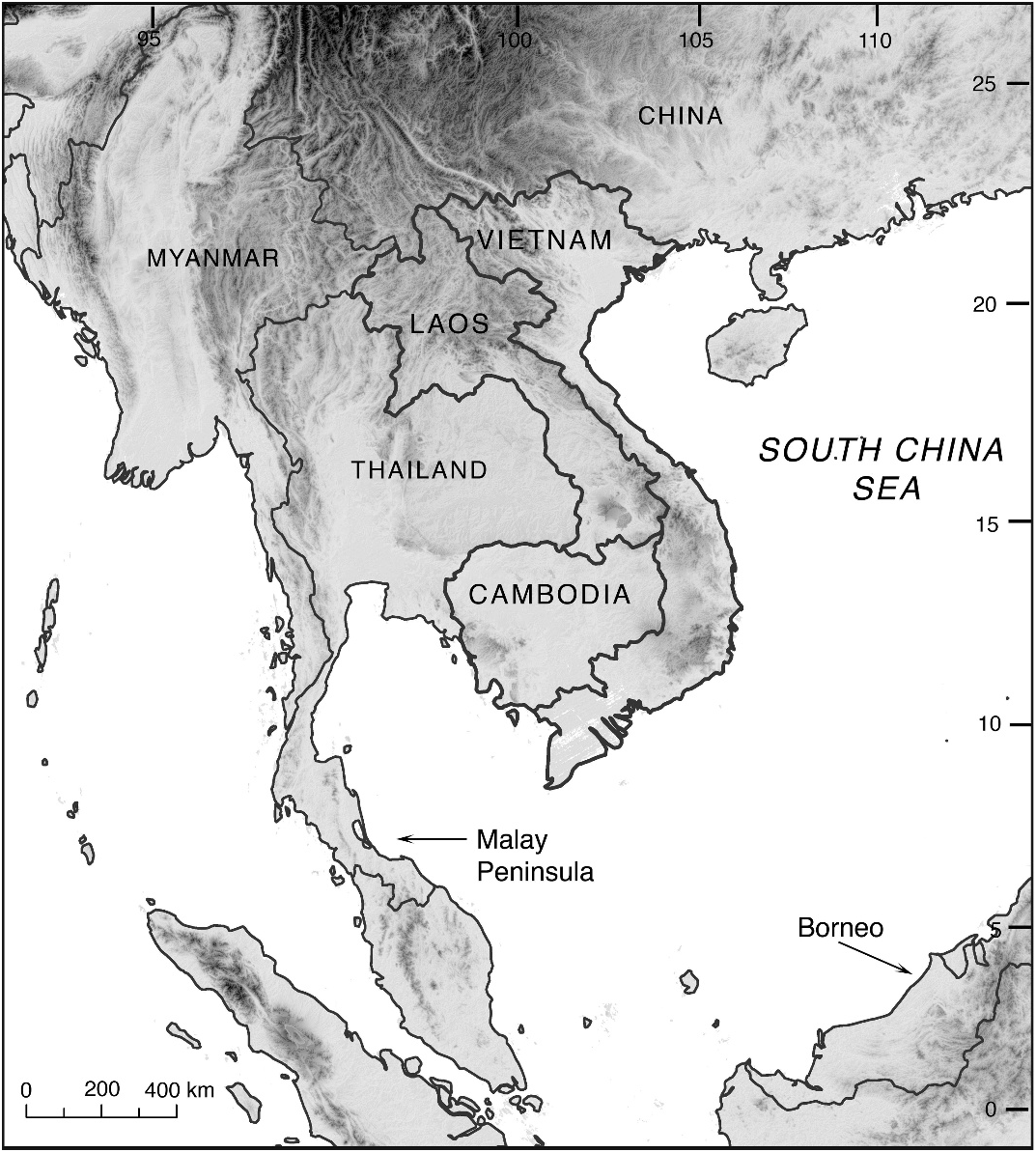
1. Variation in dispersal opportunities between northern Indochina and adjacent regions. Due to topographic differences, northwest Indochina has more limited opportunities for faunal exchange with the rest of Asia than does the northeast. Most regions adjacent to the Northwest Uplands (southern Himalayas, northeastern Myanmar and Thailand, southwestern Yunnan Province) lie at much higher elevations than northern Indochina. This may limit the dispersal of organisms both into and out of northwestern Indochina that are intolerant of altitudes exceeding ca. 3500 m. In addition, these uplands lying west of Indochina largely run north to south, which might further impede eastward dispersal into Indochina. Since the uplift of the Himalayas, and the Tibetan Plateau in particular, the dispersal corridor connecting south Asia to northwest Indochina has been restricted to a narrow band of habitat lying between the lowlands of Bangladesh and the upland areas of eastern India ( An et al., 2001).
In contrast, the Northeast Uplands and mainland southeast China are connected via montane regions lying at ca. 1000–2000 m as well as lower elevation corridors along the coast. Furthermore, during periods of lower sea level, eastern Indochina was contiguous with large areas of southeastern China (including Hainan Island) via emergent land bridges extending as far south as central Vietnam’s Hai Van pass (e.g., Lazell, 1999; Chou et al., 2001; Honda et al., 2000; Honda et al., 2001; Ota et al., 2002; see also Stuart and Parham, 2004; Yu et al., 2007a). It is unclear what role dispersal across historical land bridges with southeastern China has in structuring diversity outside the Northeast Uplands. If this were an important dispersal route for herpetofaunal lineages from Hainan Island and southeast China to eastern Indochina, then we would predict that northern Annamite populations would be genetically less divergent from those of the Northeast Uplands (and possibly southeast China to Hainan Island) than from those of the Northwest Uplands.
These geographic and paleoclimatic factors favor a greater number and frequency of dispersal events between the Northeast Uplands and southeastern China than between the Northwest Uplands and its surrounding regions, including southwestern China. The high incidence of Mainland Southeast Asian species in the Northwest Uplands may reflect limited dispersal with other adjacent regions to the north and west, while the higher biogeographic affinity between the Northeast Uplands and South China may reflect a more frequent interchange of fauna. Additionally, these factors may explain the higher species richness observed in the Northeast Uplands when compared to the Northwest Uplands.
2. Variation in climate. A second hypothesis explaining the observed patterns of species richness, divergence, and similarity in northern Indochina’s uplands is climatic variation across the region. Although the complex topography in the north makes local climates extraordinarily variable, northeastern Indochina and adjacent South China are generally both wetter and warmer than northwestern Indochina and its adjacent regions. In general, warmer and wetter climates favor many herpetofaunal lineages (including freshwater turtles, aquatic or semiaquatic snakes and lizards, and almost all amphibian species), and this hypothesis is consistent with observed greater species richness in the Northeast than the Northwest Uplands for all herpetofaunal groups except for the lizards, which are the least environmentally sensitive group. Similarly, the high variability in rainfall due to topography would result in significant geographic species restrictions for rainfall-dependent taxa. Comparative ecological analyses of the climatic and habitat requirements of potentially informative species groups can be used to further test this hypothesis. However, the detailed ecological data required is not available for the vast majority of Indochinese amphibians and reptiles. Alternatively, ecological niche modeling could be applied to determine range delimitations of species suites currently restricted to either side of the Red River in northern Vietnam (e.g., Graham et al., 2006; Raxworthy et al., 2007).
3. Red River as a dispersal barrier. A third hypothesis explaining patterns of divergence across northern Indochina’s montane uplands is that the Red River either acts as, or is coincident with, a barrier to dispersal. The Red River has a swift-flowing course through steep, narrow gorges in northern Vietnam, which has remained largely unchanged for at least the past 5 million years ( Allen et al., 1984; Replumaz et al., 2001), making it a plausible long-term barrier to dispersal movement. This theory has been advanced previously to explain the distribution patterns of a wide variety of both flora and fauna (e.g., Delacour et al., 1928; MacKinnon, 1997; Geissman et al., 2000; Orlov et al., 2001; Averyanov et al., 2003; Ziegler et al., 2008c; Zhang et al., 2009). Our results are consistent with the role of the Red River as a partial biogeographic barrier to herpetofauna.
A number of predictions can be developed to test whether the Red River is a biogeographic barrier. If the Red River does not act as a barrier to movement, one would predict an even cline in the distribution of southeast China species from east to west and southwest China /northeast Myanmar – Thailand species in the opposite direction. Our results show that 81 % (42/52) of the southeast China species found in Indochina are present in northwestern Indochina. They further show that 54 % (47/87) of the southwest Yunnan / northeast Thailand and Myanmar species found in Indochina are present in northeastern Indochina. This suggests that if there is a barrier between the two subregions, it is relatively weak with respect to adjacent faunas.
However, recent phylogenetic evidence suggests that it may play a stronger role than suggested by the above results. Genetic analyses of lineages present on both sides of the river can be used to test its effectiveness as a dispersal barrier. If the Red River is a barrier, then there should be greater genetic divergence across it than within northern Indochina’s uplands on each side of the river. If it is not a barrier (or a relatively weak one), then genetic divergence among northern Indochina’s upland populations will be strongly correlated with distance regardless of their location relative to the river’s course. The former hypothesis is supported by several phylogenetic analyses that show deep maternal lineage divergence between taxa from either side of the Red River: Paramesotriton ( Chan et al., 2001; Lü et al., 2004), Leptobrachium (Rao and Wilkinson, 2008; Zheng et al., 2008), Odorrana ( Bain et al., 2009c) , and Acanthosaura (Kalyabina-Hauf et al., 2004) .
A number of additional recent phylogenetic analyses also indicate greater affinity of northeastern Indochinese lineages to South China than to the more proximal northwestern Indochina lineages, supporting the hypothesis that dispersal across the Red River is limited. The megophryid frog Leptobrachium ‘‘ chapaense ’’ from the Northeast Uplands is sister to L. hainanense from Hainan and more distant from L. chapaensis , sensu stricto, from the Northwest Uplands (Zheng et al., 2008). Similarly, lineages of the tree vipers Viridovipera stejnegeri and Cryptelytrops albolabris from the Northeast Uplands are more closely related to those from eastern China than to the rest of Indochina’s populations (Malhotra and Thorpe, 2004a, b). This pattern of divergence between a northeastern Indochinese–southern Chinese fauna and northwestern Indochinese one is also observed in the three northernmost species of the crested gibbon genus Nomascus . The two northeastern species, N. hainanus , restricted to Hainan Island, and N. nasutus , found in northeastern Vietnam and small regions of southeast China, form a clade and are distantly related to the species found directly west of the Red River, N. concolor , a pattern that has been attributed in part to the Red River acting as a dispersal barrier ( Geissmann et al., 2000, Thinh et al., 2010).
If the Red River is a barrier to dispersal it could shape distribution patterns through a number of processes. Ancestral populations may have been split by the Red River in a vicariant event and subsequently diverged from each other. Under this mechanism, divergence times should cluster around 5 Mya or earlier. Alternatively, ancestral populations may have dispersed across the Red River after its formation and diverged in allopatry. Under these circumstances, we expect divergence times to be younger than the formation of the Red River’s current course. Finally, it is possible that these populations shared common ancestors at the river’s headwaters in Yunnan Province, China. This origin would be supported by the presence of a zone of hybridization. As with the second scenario, divergence times should be more recent than 5 Mya. All three of these processes may have occurred; relative divergence times will indicate which scenarios have shaped the observed patterns.
It is also possible that the Red River is not itself a dispersal barrier but rather coincides with a separate feature driving divergence between northwestern and northeastern Indochina. Northeastern Vietnam is part of the South China Platform whereas the remaining areas of Indochina are formed from the Indochina plate. The suture zone between these two plates lies parallel to and slightly south of the Red River. If the divergence observed between the northern Indochinese uplands were due to differences in faunal inputs between the two plates, then the suture zone itself would form a barrier coincident with the Red River. We believe this to be an unlikely explanation for subregional divergence because the collision between the plates occurred at least 65 Mya and was followed by ample opportunities for the dispersal and divergence of fauna carried on the two platforms. This hypothesis can be tested by sampling herpetofauna from the thin upland strip of the South China plate currently lying southwest of the Red River. If the hypothesis is true, this fauna should have greater faunal and genetic affinities with the Northeast Uplands than with the rest of the Northwest Uplands.
THE ANNAMITES, SOUTHERN LAO UP- LANDS, AND CENTRAL–SOUTH VIETNAM LOWLANDS: The Annamite Range, a series of adjacent uplands and plateaus with intervening lower-lying regions, lies entirely within Indochina. At its northern limit it is adjacent to the Northwest Uplands, Northeast Lowlands, and Upper Mekong Lowlands. In the south it terminates among the lowlands of eastern Cambodia and southern Vietnam. Overall, both the region and its associated lowlands have high levels of herpetofaunal species richness and endemism (tables 4, 5). The relatively depauperate nature of the Southern Annamites and Southern Lao Uplands may be attributable to the fact that they are under-surveyed per unit area compared to the rest of Indochina’s subregions.
Elevated numbers of endemic species have also been recorded from the Annamites for a wide range of other taxa (MacKinnon, 1997; Stattersfield et al., 1998; Baltzer et al., 2001; Sterling et al., 2006): at least two of the recently described ungulates ( Pseudoryx nghetinhensis , Muntiacus vuquangensis ); the recently described rabbit Nesolagus timminsi ; the doucs ( Pygathrix spp. ), a group of three leaf monkey species; and two of the three recently described babblers ( Garrulax konkakinhensis , G. ngoclinhensis ) are currently understood to be endemic to the Annamites.. Two conifers are endemic to the mountain range as well: Pinus krempfi is restricted to the Southern Annamites and the conifer P. dalatensis is restricted to the Central Annamites and Southern Annamites with the exception of a single population in the Northern Annamites in Laos (Nguyen et al., 2004; Thomas et al., 2007). Orchid endemism is also high in the Annamites, with 8 % –10 % of species in the Vietnamese sections of the central and southern sections of the range endemic to the country ( Averyanov and Averyanov, 2003; Averyanov et al., 2003).
The mechanisms responsible for high levels of Annamite endemism remain unclear and have only recently become the subject of phylogenetic work. Phylogenetic analyses of the rabbit N. timminsi and its only known congener, N. netscheri , which is restricted to the uplands of Sumatra, suggest that the species diverged ca. 8 Mya, when lowered sea levels allowed the Greater Sundas to communicate directly with southern Indochina via emergent landbridges (Surridge et al., 1999). Presumably, the ancestral Nesolagus was distributed over parts of this region during periods of lower sea levels, which allowed it to disperse between Mainland Southeast Asia and the Sundas. The occurrence of disjunct Annamite-Sundaic distributions of the snakes, Sibynophis melanocephalus and Calamaria lovii (the latter a small, burrowing species), suggests similar scenarios associated with dispersal across land bridges and subsequent isolation resulting in relict populations of these evergreen forest-dependent species. Complete regional samplings of a variety of herpetofaunal lineages will be necessary before the potential role of dispersal and isolation between mainland and insular Southeast Asia underlying the Annamites’s high proportion of endemic species can be addressed.
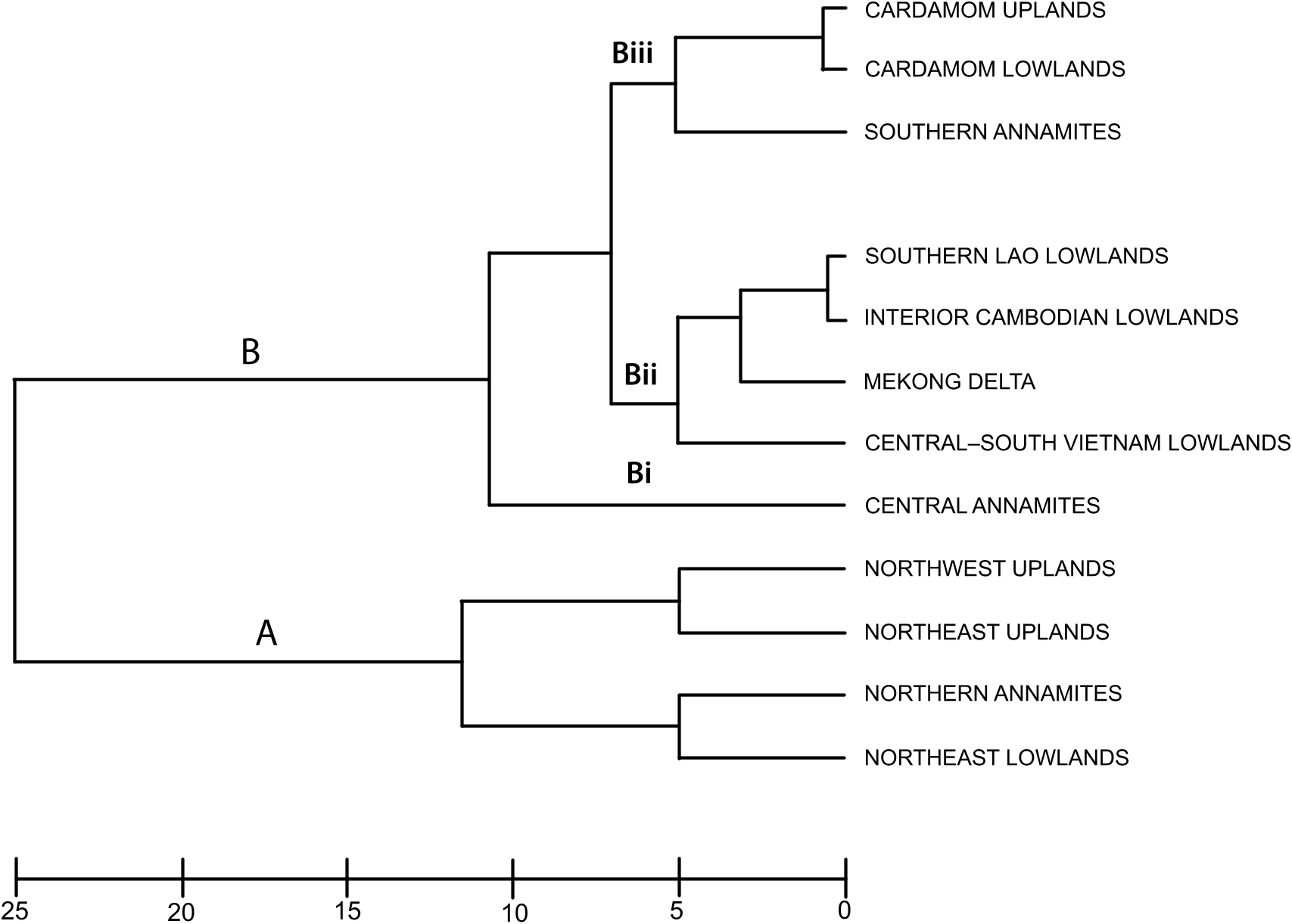
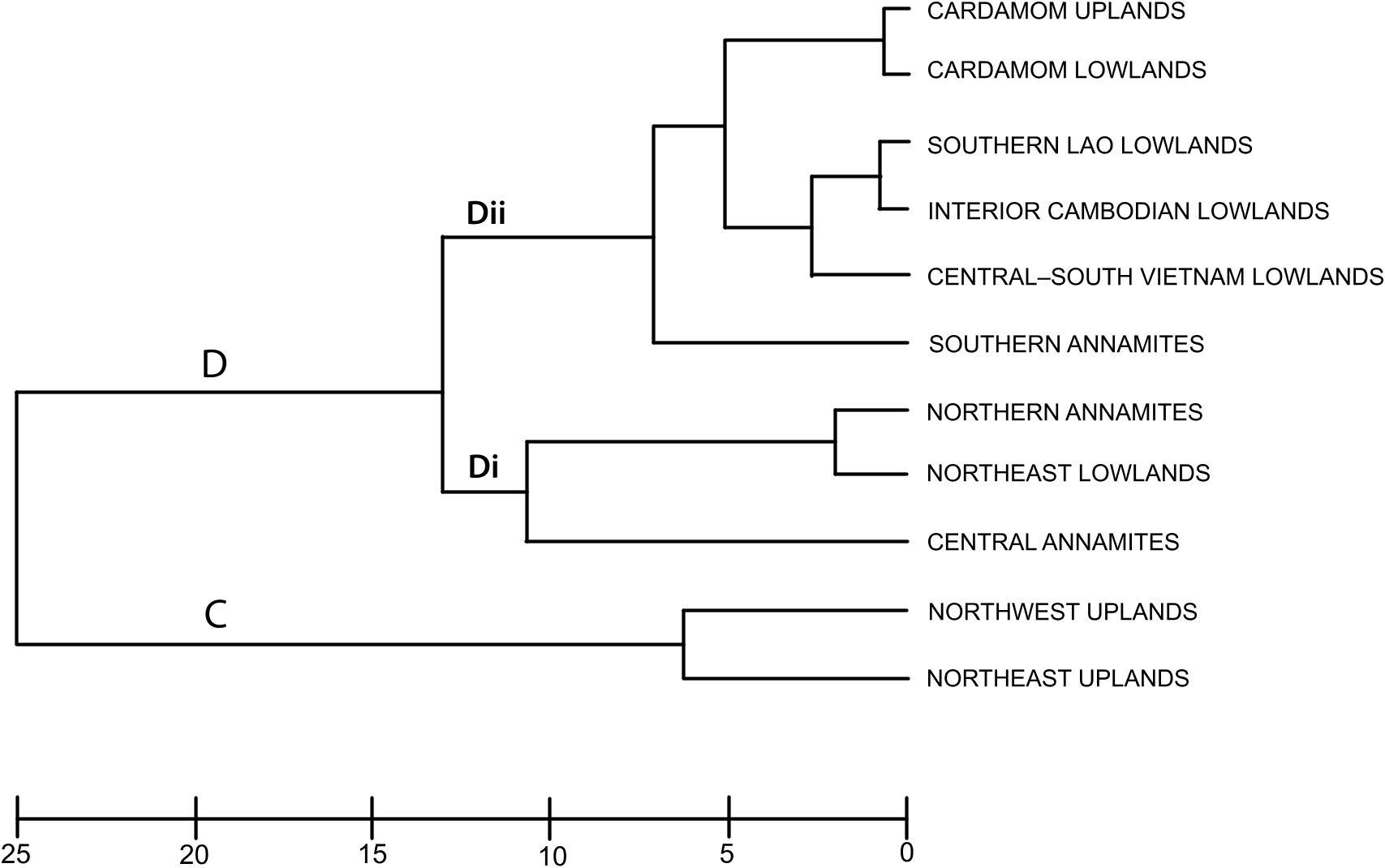
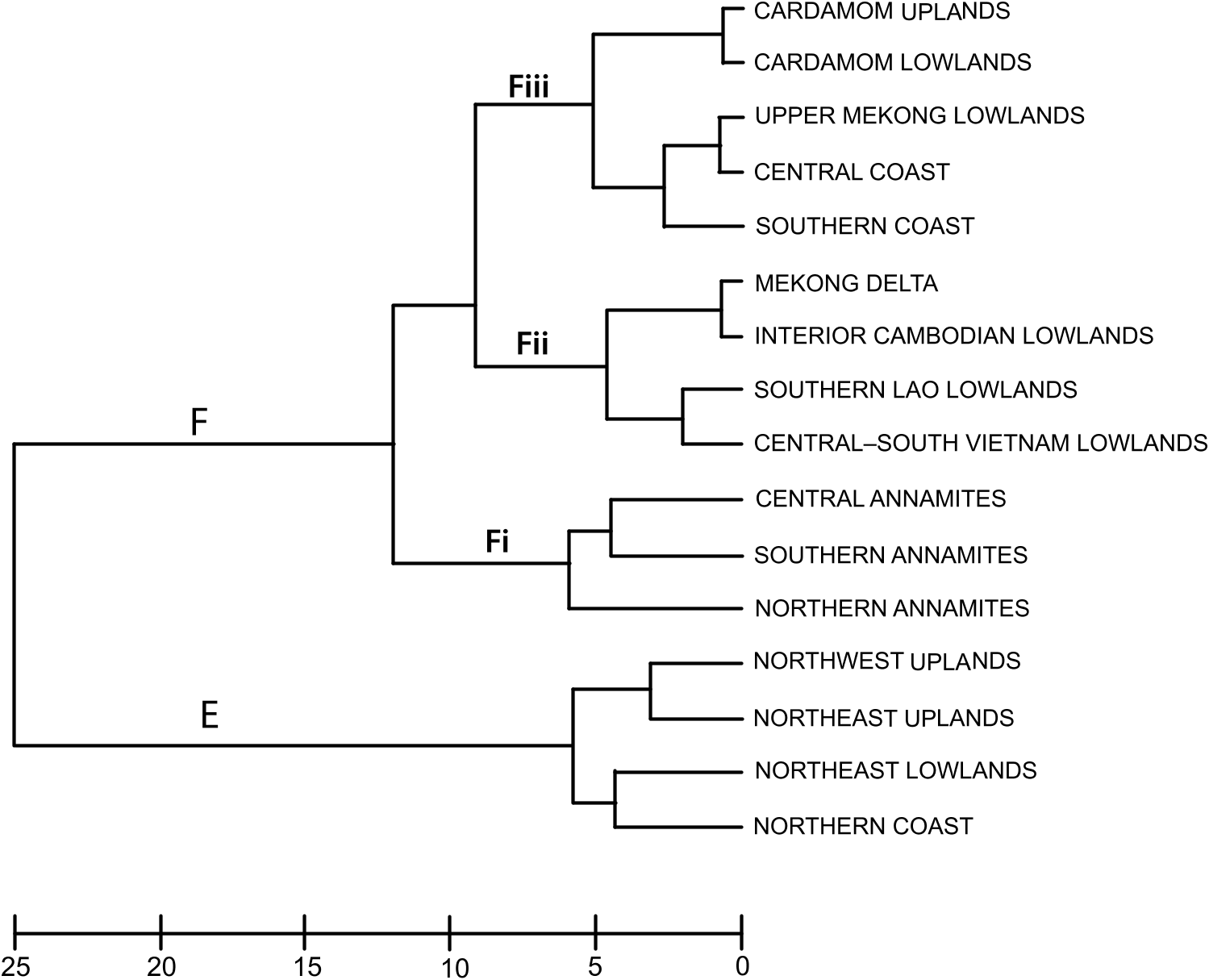
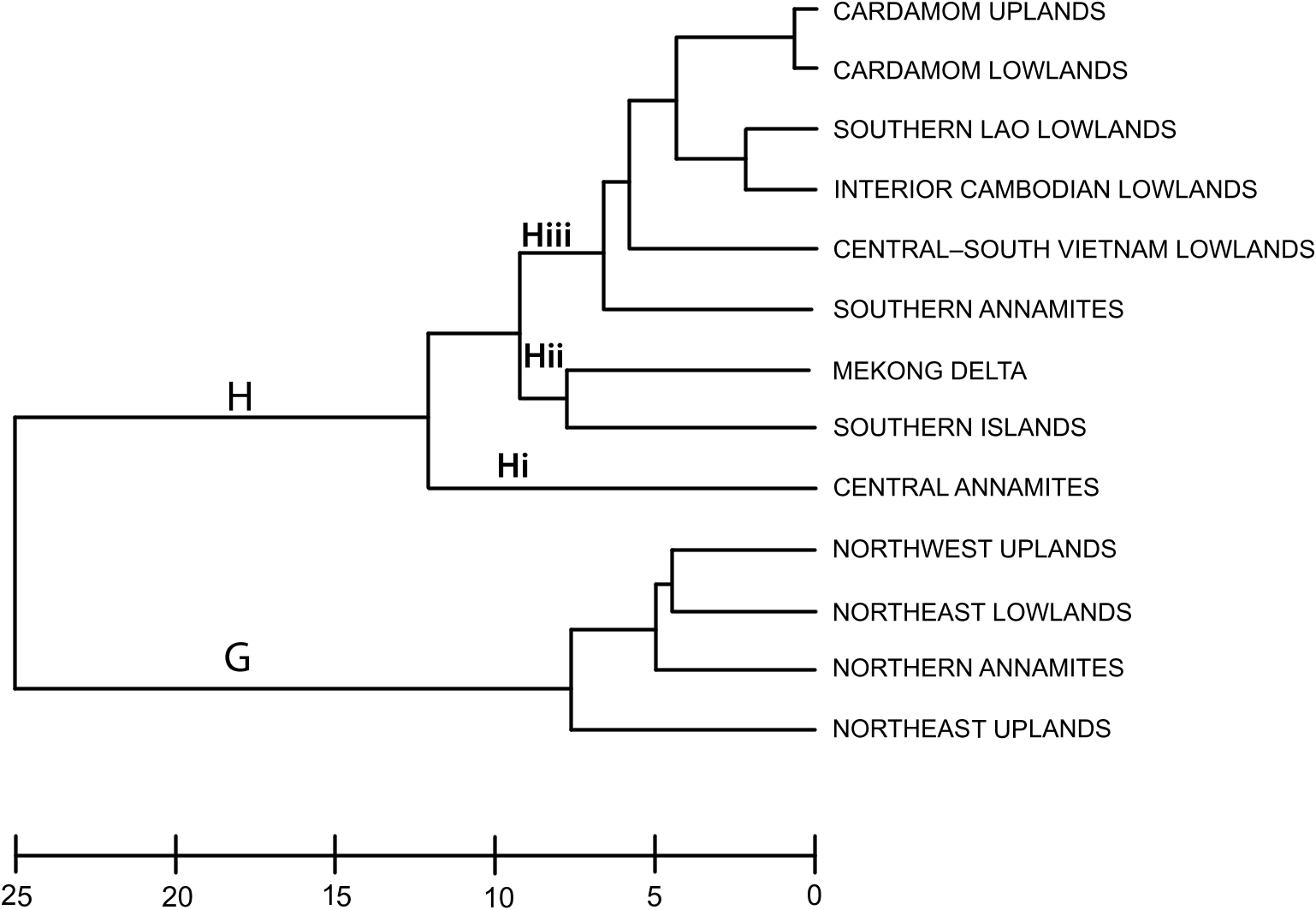
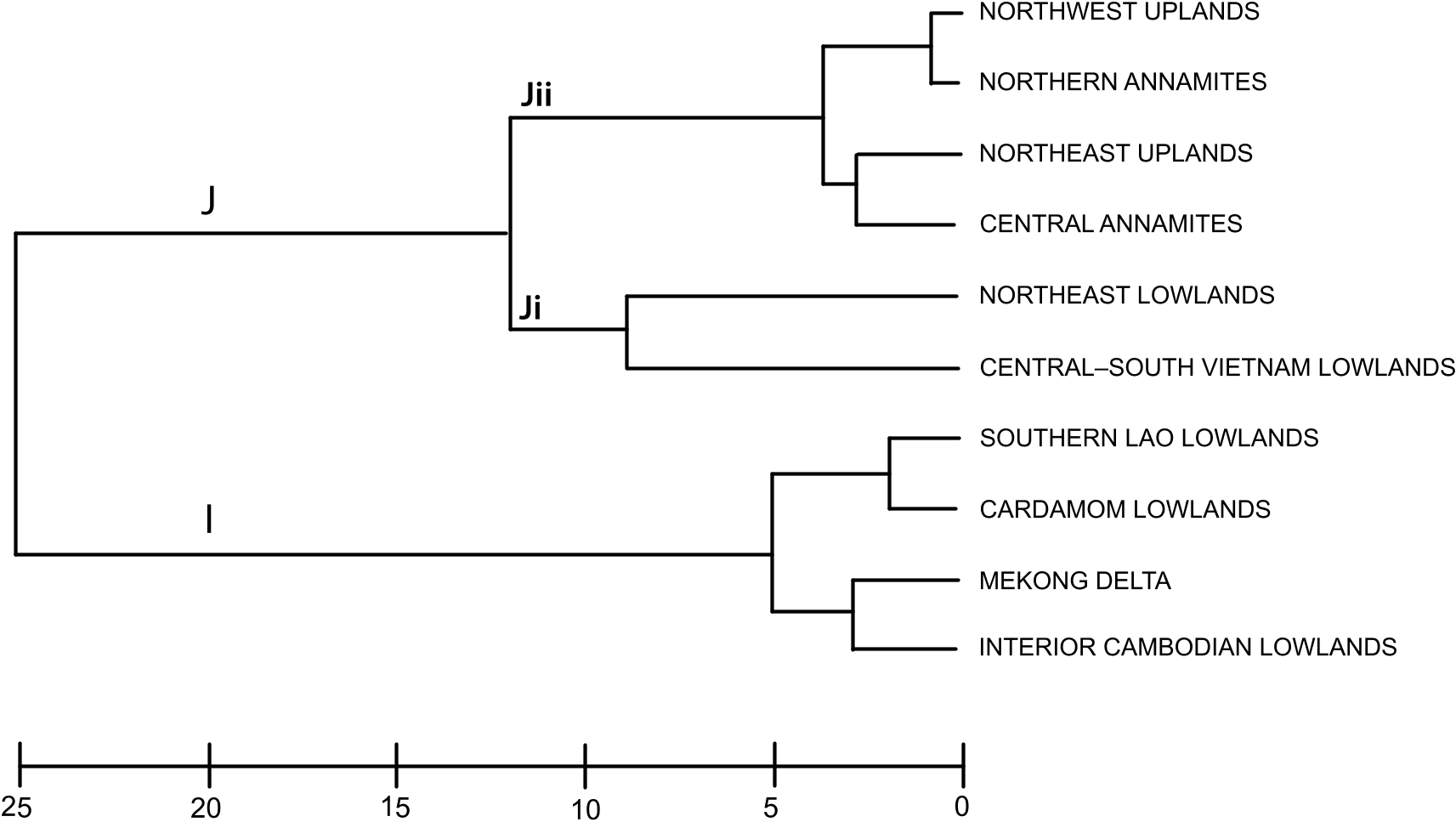
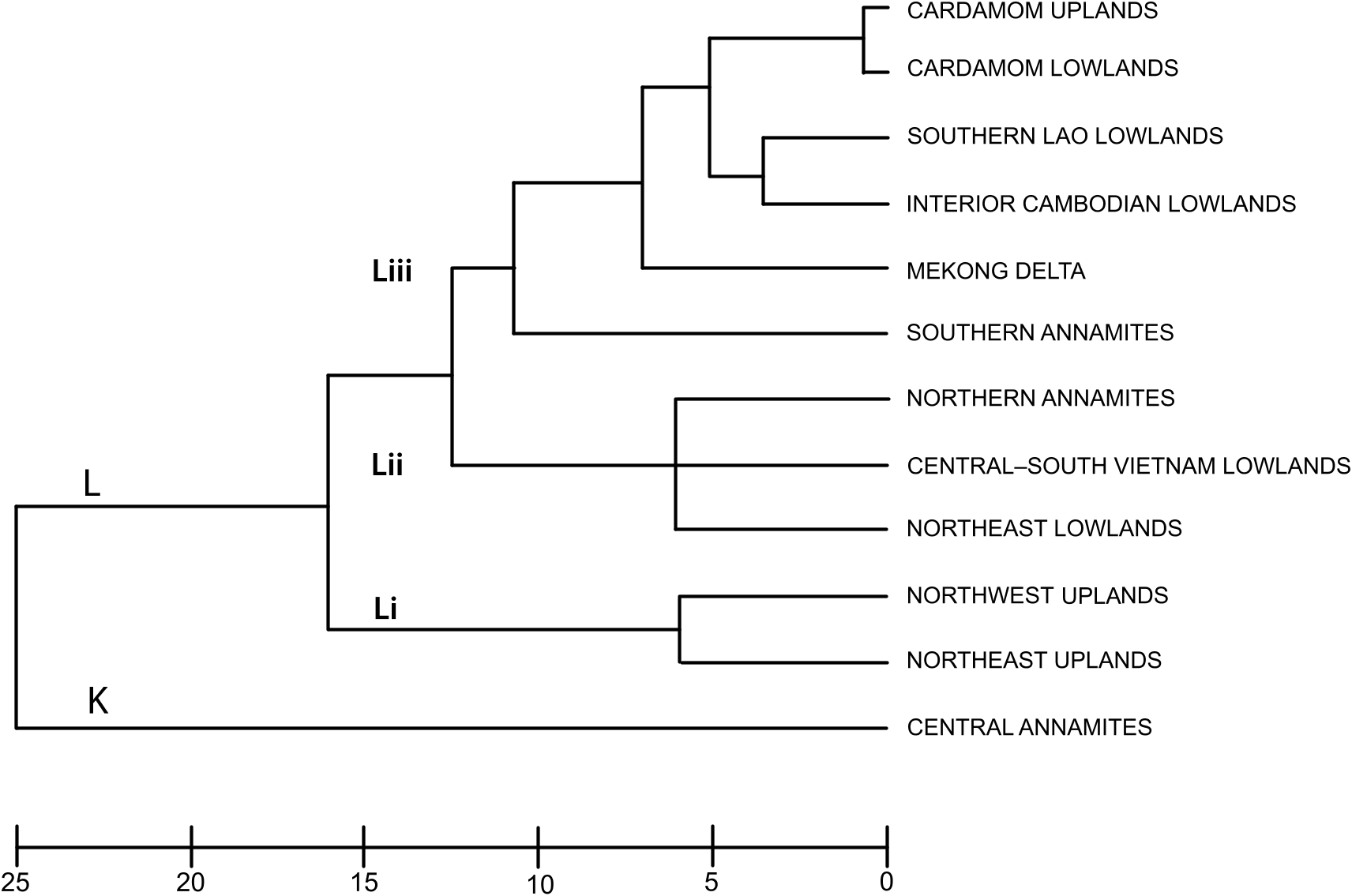
Annamite substructuring: The recognition of three distinct subregions (northern, central, and southern) within the range is supported by their strong divergence in faunal composition: of all the Annamite herpetofaunas, only the snake faunas cluster all three subregions together in the biotic affinity analyses ( figs. 4– 8 View Fig View Fig View Fig View Fig View Fig ). This contrasts with the northeast and northwest upland subregional faunas, which co-occur in the same cluster. Difference in endemic species composition is one of the main factors underlying this divergence: 76 % of the 106 Indochinese endemic species recorded from the Annamites are found within only one of the Annamite subregions. This is reflected in the cluster analyses of endemic species as all three upland subregions are placed in separate clusters ( fig. 9 View Fig ). Given the relatively small size of the Annamite uplands (115,000 km 2 total; northern: 47,000 km 2; central: 42,000 km 2; southern: 26,500 km 2), both the overall rate of endemism and the extent of substructuring within the range are notable.
Evidence for substructuring of the endemic Annamite fauna has also been recorded for other taxa. Among avifauna there is notable divergence between restricted-range species composition of the adjacent Kon Tum (Central Annamites) and Da Lat (Southern Annamites) Endemic Bird Areas (EBAs; regions covering the global ranges of two or more restricted-range bird species) (Stattersfield et al., 1998; Tordoff, 2002). The former contains nine restricted-range bird species and the latter 11, only five of which are found in both subregions ( Rheinardia ocellata , Garrulax milleti , G. vassali , Jabouilleia danjoui , and Sitta solangiae ) (Tordoff, 2002; Ounekham and Inthapatha, 2003; Collar, 2006). Among primates, the red-shanked douc ( Pygathrix nemaeus nemaeus ) occupies the Northern Annamites and Central Annamites whereas the black-shanked douc ( P. nigripes ) is concentrated largely in the Southern Annamites and associated lowlands; the intermediate gray-shanked (P. n. cinerea) form lives in the southern section of the Central Annamites, overlapping both of the other taxa. The Saola ( Pseudoryx nghentinhensis ) is restricted to the northern and Central Annamites. The distribution of the additional recently described large mammals in the Annamites remains unclear due to a long history of exploitation and open phylogenetic questions ( Amato et al., 2000; Groves and Schaller, 2000).
The presence of at least three distinct biogeographic units in the Annamite Range is also concordant with the patterns described in some recent phylogenetic studies. The freshwater turtle Cuora galbinifrons species group was found to have three distinct lineages, with a southern Annamite clade sister to a central plus northern Annamite clade (although the provenance of the southern Annamite clade was inferred from trade data) (Stuart and Parham, 2004, and references therein). Phylogenetic analyses of primates are concurrent with this pattern. A phylogeny of the douc complex indicates a sister relationship between P. n. nemaeus (northern and Central Annamites) and P. n. cinerea (Central Annamites), with P. n. nigripes (Southern Annamites) sister to this clade (Roos and Nadler, 2001).
Northern Annamites: Indochina’s Northern Annamites is contiguous with both the Northwest Uplands to the north and the Central Annamites to the south. Its geographic location between the northern uplands and the remaining Annamite uplands subregions is reflected in their faunal composition and biotic affinity. The Northern Annamites ranks fourth in overall species richness and the cluster analyses suggest greater faunal overlap with adjacent northern subregions than southern ones: of all groups, only the snakes cluster tightly with a southern subregion, the Central Annamites ( figs. 4–8 View Fig View Fig View Fig View Fig View Fig ). The subregion’s endemic species cluster with both the Northeast Lowlands and the Central–South Vietnam Lowlands in the biotic affinity analyses of endemic species ( fig. 9 View Fig ). Of the 32 species of endemic Indochinese fauna recorded from the Northern Annamites, half are strictly northern Indochinese endemics and half are known from both northern and southern Indochina. This contrasts with the Northeast Uplands and Northwest Uplands, which are dominated by strictly northern Indochinese endemics, and the Central and Southern Annamites, which are dominated by strictly southern endemics. Biogeographically, the Northern Annamites also appear to be transitional between northern and southern Indochina. The proportion of South China fauna present in the Northern Annamites (14 %) is intermediate between the high values recorded in the northern uplands (24–31 %) and the much lower ones in the Annamite uplands to the south (5–9 %). Given the above results we hypothesize (1) that northern Annamite populations of a given species should be less divergent from populations in other upland subregions than are populations between other upland subregions, and (2) that, given their clustering with northern subregions, Northern Annamite populations should have lower levels of genetic divergence from the Northern uplands than from the Central and Southern Annamites.
Central Annamites: The central Annamite uplands form a contiguous montane block with the Northern Annamites; however, it is separated from the Southern Annamites by low-lying areas covered by a mixture of evergreen, semievergreen, and dry forest types. The Central Annamites ranks third among Indochinese subregions in total species richness. Central Annamite herpetofauna as a whole forms a unique group within a southern cluster of subregions ( fig. 4 View Fig ), a pattern that is also true for the lizard fauna ( fig. 7 View Fig ). The other three taxonomic groups exhibit greater variation in their affinity with other subregions and cluster with at least one northern subregion: the Central Annamites amphibian fauna clusters with the adjacent Northern Annamites + Northeast Lowlands ( fig. 5 View Fig ); the Central Annamites snake fauna is part of a strictly Annamitic cluster ( fig. 6 View Fig ); and turtles are the only Central Annamites fauna clustering most closely with a nonadjacent subregion, the Northeast Uplands ( fig. 8 View Fig ). The Central Annamites has the highest level of herpetofaunal endemism in Indochina, forming a unique cluster in the affinity analysis; this indicates that its composition is highly divergent from the rest of Indochina ( fig. 9 View Fig ).
Because of its intermediate position between the northern and southern upland regions of Indochina, previous researchers have proposed that the Central Annamites serves as a refuge for northern upland herpetofauna whose ecological niches require cooler, humid, upland habitats (e.g., de Laubenfels, 1975; Brandon- Jones, 1996; Inger, 1999; Surridge et al., 1999; Orlov, 2005). We detected 113 species from the central Annamite fauna that were also found in the Northwest Uplands and/or the Northeast Uplands (the Northern Annamites were excluded from this analysis due to their physical continuity with the Central Annamites). Eight of these species (7 %) are restricted to regions above 300 m, and only one is restricted to areas above 800 m ( Geomyda spengleri ). A similar pattern of faunal overlap is observed for the Central Annamites and southern uplands (Southern Annamites, Southern Lao Uplands, and Cardamom Uplands). Of the 120 species shared between these regions, 9 (8 %) are restricted to regions above 300 m and only one is restricted to areas above 800 m ( Leptobrachium pullum ).
Although the numbers are relatively low, these two suites of species are notable for being entirely exclusive of each other. This suggests that the Central Annamites may capture upland-restricted species dependent on evergeen forests at both the southernmost and northernmost extent of their ranges in Indochina. The ranges of the upland Central Annamite species that are also known from the Northwest Uplands and Northeast Uplands (i.e., the frogs Leptolalax bourreti , Babina chapaensis , Rhacophorus feae , Theloderma gordonii ; the tree viper Viridovipera gumprechti ; the elapid Bungarus slowinskii ; and the turtles Platysternon megacephalum , and Geoemyda splengeri ) extend to South China and/or Mainland Southeast Asia. The ranges of Central Annamites fauna also known from the southern uplands are dominated by Indochinese endemics. Three of these species are frogs endemic to the Annamites: Brachytarsophrys intermedius , Leptobrachium mouhoti , and Chiromantis laevis ; three are Indochinese endemics: the snake Fimbrios klossi , the frog Ophryophryne gerti , and the lizard Acanthosaura capra ; two are frogs found across South China and/or Mainland Southeast Asia: the frogs L. pullum and Feihyla palpebralis ; and one snake extends to the Southern Himalayas: Calliophis intestinalis .
Biogeographical analyses mirror this pattern. There is a significantly higher proportion of South China and/or Mainland Southeast Asia species in the shared northern uplands–Central Annamites fauna (34 %) than in the shared southern uplands–central Annamite fauna (21 %) (x 2 5 7.018, df 5 1, p, 0.01). Similarly, the proportion of endemic Indochinese fauna shared between the southern uplands and the Central Annamites (17 %) is higher than that shared between the northern uplands and the Central Annamites (4 %) (x 2 5 10.854, df 5 1, p, 0.001). Both the northern and southern upland herpetofauna share an equal proportion of Pan-Asia species (63 %) with the Central Annamites.
Southern Annamites: The Southern Annamites consists of a suite of plateaus that are isolated geographically from the Central Annamites. It is also a younger formation created by different orogonies than those that uplifted the Central Annamites. Affinity analyses indicate that the combined herpetofauna of the Southern Annamites cluster most closely with the Cardamom Uplands and Caradamom Lowlands ( fig. 4 View Fig ) despite their geological uniqueness and separation from these two subregions by a large expanse of lowland delta ( fig. 1 View Fig ). This clustering likely results in large part from the relatively low number of surveys conducted in these three subregions and the underlying assumptions of the clustering algorithm that weights shared presences and absences equally (see Methods). Although all three subregions harbor more than 15 % of Indochina’s total herpetofauna, their overall species richness ranks are low (table 4). Despite this, the Southern Annamites ranked second overall in regional endemism per unit area (table 5). Further surveys of the Southern Annamites will be necessary before its species richness, endemic composition, biogeographic affinities, and relationship to the rest of Indochina’s subregions can be accurately assessed.
Southern Lao Uplands: Given its relatively large size (47,500 km 2, similar to both the Northern Annamites and Central Annamites), the presence of evergreen forests, as well as its geographic complexity and close proximity to the species-rich Central Annamites, we predict that this region should be relatively speciesrich. However it remains under-surveyed per unit area when compared to other subregions of Indochina, having been rigorously surveyed for herpetofauna only twice (Teynie et al., 2004; Stuart, 2005b). We excluded the Southern Lao Uplands from the cluster analysis for both richness and endemism because of its low overall species numbers.
A relatively high proportion of the Southern Lao Uplands herpetofauna is endemic (33 %), a level that is similar to that recorded for the Central (33 %) and Southern Annamites (25 %) and the Southern Islands (25 %). A roughly equal proportion of this endemic fauna is made up of species restricted to southern Indochina and those found in both northern and southern Indochina. A similar ratio is observed in the Central–South Vietnam Lowlands, Southern Lao Lowlands, and Interior Cambodia Lowlands; the remaining southern subregions’ endemic faunas are dominated by southern Indochinese species.
Evidence from other research suggests that the Southern Lao Uplands might be an extension of the Central Annamites rather than a separate biogeographic unit. A pattern of shared fauna between the two subregions has been suggested for butterflies (Monastyrskii, 2007) and plants ( Averyanov et al., 2003), and at least two bird species endemic to the Annamites have also been recorded from the Southern Lao Uplands ( Garrulax vassali , Macronous kelleyi ) (Tordoff, 2002; Ounekham and Inthapatha, 2003). Given the lack of data on species composition of the Southern Lao Uplands, it is unclear whether this affinity is also true for the herpetofauna. A species comparison of the Southern Lao Uplands and nearby uplands shows that there is extensive faunal overlap with the Central Annamites (81 %) and the Southern Annamites (70 %) and a lower level of exchange with the Cardamom Uplands (42 %). However, only 57 % of the Southern Lao Uplands fauna is also found on the Khorat Plateau of neighboring Thailand. These findings are congruent with those of Teynie et al. (2004), who suggested that the Bolovens Plateau of the Southern Lao Uplands might act as a refuge for herpetofauna associated with upland and highland humid areas.
The high similarity between the herpetofauna of the Bolovens Plateau and adjacent upland subregions may be attributable to its relatively recent age; a basaltic formation created ca. 800,000 years ago ( Fontaine and Workman, 1997; Attwood and Johnston, 2001). It is therefore likely that the plateau was colonized recently by fauna from neighboring upland areas unaffected by the giant lava flows that created it. This hypothesized recent colonization is congruent with at least one phylogenetic analysis, which found that a central Annamite population of Viridovipera vogeli was more closely related to populations on the Dongreak Mountains ( Thailand), than those of the intervening Bolovens Plateau (Malhotra and Thorpe, 2004a).
Central–South Vietnam Lowlands: The Central–South Vietnam Lowlands has high levels of both species richness and endemism. In general, the subregion has the greatest faunal overlap with Indochina’s other lowland subregions ( figs. 4–9 View Fig View Fig View Fig View Fig View Fig View Fig ). This is likely due to the high number of Pan-Asia species distributed across Indochina’s lowlands, although they cluster with the northern subregions and Central Annamites in our affinity analysis.
The Central–South Vietnam Lowlands stands out as the only non-island lowland subregion with a notable number of endemic species (27); it ranks fifth in endemic species richness for all major groups except lizards (table 5). This is largely due to its proximity with the adjacent Central and Southern Annamite uplands with which it shares 67 % of their endemic species. A finer-scale mapping of this subregion than is currently possible would allow us to define those areas that are most faunally similar in composition to the Central Annamites and Southern Annamites, respectively, and identify possible dispersal corridors between these upland formations as well as with the adjacent Northeast Lowlands.
It is also possible that this lowland region contains more than one biogeographic unit. The Central–South Vietnam Lowlands is a diverse area composed of three largely noncontiguous blocks covering almost five degrees of latitude and including three major forest habitat types (evergreen, semievergreen, and deciduous dipterocarp). Although portions of the Central–South Vietnam Lowlands are adjacent to the Northeast Lowlands, they do not share as many species as do the Central–South Vietnam Lowlands and the partly adjacent Southern Lao Lowlands. This can be attributed to habitat differences and as yet unrecognized structuring of diversity within the lowland units.
THE LOWER MEKONG WATERSHED AND CARDAMOM UPLANDS AND LOWLANDS: The southernmost third of Indochina is dominat- ed by the large, ecologically and biotically uniform lower Mekong watershed, which encompasses three lowland subregions: the Southern Lao Lowlands, Mekong Delta, and Interior Cambodia Lowlands. The species richness of these lowlands is high, though its herpetofauna is dominated by widespread, Pan-Asia species. There is little evidence of substructure in the distribution of amphibians and reptiles within the region. In contrast to Indochina’s other lowland areas, the lower Mekong watershed has few adjacent upland regions and a low, uniform elevation except for a series of low parallel rocky hills separated by agricultural plains in its southern regions.
The only portion of Cambodia lying outside the Mekong watershed is the Cardamom Massif. The Cardamom Uplands and its associated Lowlands share over half of their nonendemic species (61 %), although of the 11 endemic species found in the two subregions, only two occupy both the uplands and lowlands. Because of their proximity to Thailand, both subregions are dominated by species also found in Thailand. A large number of these are species restricted to Mainland Southeast Asia, a fauna that makes up 19 % of upland and 16 % of its lowland species richness. Overall, the Cardamoms are relatively depauperate of herpetofauna: the uplands rank fourth from last and the lowlands last in species richness and endemism. Recent bird surveys in the Cardamoms also indicate a relatively depauperate avifauna compared to other Indochinese uplands, although three restricted-range species are found there, including one endemic to the area ( Garrulax ferrarius ) ( Eames et al., 2002; Collar, 2006). This depauperate fauna may be partly attributable to low survey efforts. The Cardamoms are relatively under-surveyed per unit area compared to the other subregions in Indochina; and to date there have been only four major surveys specifically targeting the Cardamoms ( Daltry and Chheang, 2000; Daltry and Wüster, 2002; Ohler et al., 2002; Stuart and Emmett, 2006; Grismer et al., 2007a, 2008a, b) supplemented by short site-specific visits and patchy survey efforts (e.g., Mouhot, 1864; Smith, 1928; Bauer and Das, 1998; Platt et al., 2003, 2006; Holloway and Sovannara, 2004).
Studies of mammalian distribution patterns, and of primates in particular, have supported the hypothesis that the Mekong River presents a geographic barrier to dispersal ( Fooden, 1996; Nadler et al., 2005; Meijaard and Groves, 2006). We found little evidence of this for amphibians and reptiles. The three southern subregions through which the Mekong River flows (Mekong Delta, Interior Cambodia Lowlands, Southern Lao Lowlands) clustered closely together both for total and endemic species richness. Furthermore, over half of Indochina’s total herpetofauna (256 species) are also found in Thailand, which is largely separated from Indochina by the Mekong River. The weak role of the Mekong as a biogeographic barrier is congruent with phylogenetic analyses of Viridovipera vogeli , whose populations from the Central Annamites (west of the Mekong) are more closely related to Thai populations from the Dongraek Mountains (east of the Mekong) than they are to Boloven populations (west of the Mekong) (Malhotra and Thorpe, 2004a). Furthermore, the monotypic Mantheuys phuwuanensis , known from the Upper Mekong Lowlands and adjacent Thailand is found on either side of the Mekong River.
Historically, there has been little opportunity for populations on either side of the Mekong to diverge. The river has undergone a complex series of captures and course shifts over the past 7 million years (Rainboth, 1996). Until the late Pleistocene its upper section flowed westward around the Khorat Plateau and into the Gulf of Thailand via the Chao Phraya River, whereas the lower Mekong south of Khong Island (located just north of the Lao- Cambodian border) flowed westward, entering the gulf near Kampot on Cambodia’s southern coast, until ca. 5000 years ago ( Attwood and Johnston, 2001). Today it flows to the east of the Khorat Plateau and enters the gulf in southern Vietnam. Given these recent shifts and captures, the lower Mekong Basin of Indochina may have formed a contiguous region until quite recently, allowing little time for divergence of either genetic or faunal composition. This is in contrast to the Red River, which has followed its current course for a minimum of 5 million years.
Taken together, the analyses and history of the Mekong River’s course shifting suggest that the Mekong Delta, Interior Cambodia Lowlands, and Southern Lao Lowlands should be a considered a single biogeographic unit instead of three subregions divided by political boundaries. Considering the lower Mekong watershed as a single unit would create an enormous subregion, the largest in Indochina, covering approximately onequarter of Indochina’s total land area with an overall species richness of 213 (ranked 8 out of 10, controlled for area), 34 of which are endemic species (ranked 7 out of 10, controlled for area). Although this extensive area’s fauna is composed primarily of Pan- Asia species (62 %), it holds 14 freshwater turtles, representing over half of Indochina’s overall freshwater turtle biodiversity. The same number of turtles occurs in the Northeast Lowlands, but only four species are common to both areas ( Cuora mouhotii , Cyclemys oldhami , Indotestudo elongata , and Pelochelys cantorii ).
Recent survey results suggest that there are still undiscovered species in the isolated hills and outcroppings in the lower Mekong watershed as well as nearshore islands that would have been connected to the mainland during glaciation events. Isolated rocky outcrops within the lower Mekong Delta have been shown to house endemics of the gekkonid genus Cnemaspis (a genus known from the Sundas, Malay Peninsula, Mainland Southeast Asia, and southern regions of Cambodia and Vietnam). Until very recently, there was a large gap in the distribution of Cnemaspis between southeast Thailand ( C. chanthaburiensis ) and the Con Dao Islands in the South China Sea ( C. boulengeri ). Cnemaspis chanthaburiensis has since been recorded from the Cardamoms and six more endemic congeners described from the Cardamoms and southern Vietnam, including small rocky hills of the Mekong Delta and offshore islands of Rach Gia Bay, which represent partly submerged areas of the Cardamom Massif ( Grismer et al., 2010 a, b). The endemic rupicolous pitviper, Cryptelytrops honsonensis , has also been described from Rach Gia Bay ( Grismer et al., 2008c). Grismer and Ngo (2007) further point out that these lower Mekong outcroppings would have existed as islands during sea level highstands, greatly reducing any opportunity for genetic exchange. They postulate that if Indochinese Cnemsapis species form a monophyletic group, it is likely that upland isolation due to sea level highstands drove speciation, although the timing of these highstands has yet to be calibrated. These processes may also explain the geographic distribution of C. honsonensis .
COASTS AND ISLANDS: The unique attributes of Indochina’s coastal and island subregions strongly structure their faunal composition. Indochina’s narrow coastal subregions span almost its entire latitudinal extent and present relatively harsh conditions, particularly to amphibians: the deltas are brackish near the coasts and elsewhere conditions are largely arid. Only nine of the region’s 120 coastal species are amphibians, representing less than 5 % of Indochina’s total amphibian fauna. The coasts are also Indochina’s most faunally depauperate subregions; snakes are the only major taxonomic group with a sizable presence in all three coastal areas where they make up over half of the herpetofauna. Although historically present in the Central Coast and Southern Coast, Crocodylus porosus , one of two species of crocodiles known from Indochina, has now become virtually unknown in these areas. Two-thirds of the coastal herpetofauna are Pan-Asia species (67 %) and, as a whole, the fauna has broad ecological tolerances: 58 % of all species are found in open habitats (including those also found in forests) and 57 % can be found at all elevations. However, these coastal regions are also home to some unique endemic species not found elsewhere in Indochina. This includes a suite of reptiles known only from the southern coasts: two colubrid snakes, Oligodon macrurus and O. moricei ; two gekkoes, Dixonius aaronbaueri and Gekko grossmanni ; and one agamid lizard, Leiolepis ngovantri .
The island subregions, as defined in this analysis, remain somewhat artificial units because they combine survey data from individual islands, which may be poorly comparable. We would have preferred to group islands by geological and geographic relatedness and treat each group as a separate subregion. Regardless, there are some general comments that we can make about the richness and distribution of Indochina’s island fauna.
As with the coasts, the species composition of Indochina’s island amphibian and reptile communities appear to be strongly affected by local climate and habitat. Aridtolerant snakes and lizards are the only groups to have significant levels of richness on the offshore islands and of the 17 Indochinese endemic species found on islands, 15 are lizards. The remaining two endemics are the viper Cryptelytrops honsonensis and the frog Limnonectes dabanus . In contrast, no turtles have been recorded from the offshore islands and among the eight amphibians present, six are widespread, ecologically tolerant Pan-Asia species. All islands lack the permanent and diverse freshwater habitats found on the mainland, with the single exception of the Rach Cua Can River on Phu Quoc. This distribution pattern is similar to that recorded in the generally dry coastal areas where snakes and lizards also have higher species richness than the more freshwater-dependent amphibians and turtles. However, only two Indochinese endemics known from the coasts are also found on the islands: the parthenogenetic lizard Leiolepis guentherpetersi known from the Central Coast and Southern Islands, and L. guttata from the Southern Coast and Northern Islands (the unusual distribution pattern of the latter species suggests that identification of this taxa from the Northern Islands needs to be revisited).
Indochina’s island faunas largely resemble relict mainland faunas, reflecting their likely origins as remnant communities of the Sunda Shelf, which were last connected to the mainland approximately 15,000 years ago. This is supported by the suite of Indochinese endemic species known from the Southern Islands and interior mainland, but not from the intervening coasts: the skink Sphenomorphous rufocaudatus and the gekko Hemidactylus vietnamensis (both widespread throughout Indochina across elevations); the fossorial lizard Dibamus montanus and the frog Limnonectes dabanus (both known only from southern Indochina); and the gekko Cyrtodactylus paradoxus (also known from the Mekong Delta). Most of the islands lie close to the mainland, the major exception being Con Dao, a small granitic and ruggedly mountainous island archipelago 80 km southeast of the Mekong Delta (Le, 1989; BirdLife International in Indochina and MARD, 2004). Its greater isolation during periods of sea level fluctuation likely explains the presence of three lizard species endemic to the archipelago ( Cnemaspis boulengeri , Cyrtodactylus condorensis , and Dibamus kon- daoensis) ( Darevsky and Szczerbak, 1997; Darevsky, 1999; Honda et al., 2001). The remaining seven island endemics (i.e., not known from the mainland) are known from geological extensions of the Cardamom Massif: the rupicolous viper Cryptelytrops honsonensis , and a suite of gekkoes scattered among Phu Quoc and small islands of Rach Gia Bay (see lower Mekong watershed section above). Nonherpetofaunal taxa endemic to this archipelago include two squirrels ( Ratufa bicol- or condorensis , Callosciurus finlaysonii germaini ) ( Dao, 1967; van Peenen et al., 1969; Kuznetsov, 2000) and a primate ( Macaca fascicularis condorensis ) ( Brandon-Jones et al., 2004).
Only a single Malaya species has been recorded from Indochina’s coasts and islands: the aquatic snake Acrochordus javanicus , which has been recorded from the Southern Coast. Widespread in southern Indochina’s lowland regions, it is the only Malaya species not restricted to forested habitats. This is consistent with the previous observation from the Annamite region that the few strictly Malaya species found in Indochina represent apparently relict populations of evergreen-forest-dependent species, which were more widely distributed during historical periods when this forest type was more widespread.
GENERAL PATTERNS OF DISTRIBUTION
Indochina’s major herpetofaunal groups share some broad habitat associations: species in all five groups occur in higher numbers in forested environments; are more likely to occupy a single habitat than multiple ones; and, with the exceptions of turtles, are more likely to be found at elevations above 300 m. However, there is also significant variation among the major taxonomic groups and between suites of species with different geographic distributions and biogeographic affinities. It should be noted that, given the active state of herpetofuanal work in Indochina, any conclusions concerning the distribution of its amphibians and reptiles are contingent on the results of further research. This is particularly true for species currently considered regional endemics, 69 % of which have been described since 1997. This strongly suggests that knowl- edge of their distribution and ecology will change over time.
The four major taxonomic groups differ with respect to microhabitat associations, the number of microhabitats used, and elevation zone. Turtles are the only major group with a large proportion of species (50 %) restricted to areas below 300 m. Amphibians are more likely than reptiles to be restricted to forests and uplands, and are the only group with a sizable suite of species (30 %) limited to areas above 800 m. Snake and lizard species occur more frequently in open areas, across wider elevation gradients, and in greater variety of microhabitats (i.e., rupiculous, fossorial) than do amphibians or turtles. Some of these patterns correlate with the broad habitat requirements of each taxonomic group. Indochina’s turtles are dependent on water resources, but a subset of species (four; 14 % of the fauna) is able to tolerate brackish, estuarine environments. Given low overall turtle species richness (28), the occurence of these species in the region’s slow lowland deltas may contribute to the group’s overall association with lowland habitats. Although the breeding ecology of Indochina’s freshwater turtles is imperfectly known, it is also possible that lowland areas are more conducive to their egg-laying habits than the oftenrocky banks of upland and highland waterways. Amphibians are dependent on freshwater resources and the vast majority requires bodies of water associated with forests. Today, this combination is found most commonly in Indochina’s extensive upland and highland areas. However, historically lowland regions would also have hosted suitable large areas of forest that could be utilized by species dependent on slower or still freshwater environments. Snakes and lizards are less freshwater-dependent and therefore capable of occupying a wider variety of habitats, such as open, rocky, or dry areas, which amphibians and turtles cannot tolerate.
ENDEMIC SPECIES: The habitats occupied by endemic species are significantly more restricted than those of nonendemic species. In general, endemic species occupy a narrower range of elevations and a subset of the microhabitats used by their nonendemic counterparts, two ecological variables that are frequently correlated (e.g., fast-flowing streams in upland environments, rupicolous niches on low-lying rocky outcrops). This negative correlation between ecological niche width and geographic range is shared by the three lineages in which it could be analyzed (amphibians, snakes, and lizards). However, within each group, the distribution (geographic and taxonomic) of endemic species is often strongly correlated with lineage-level habitat requirements. As a result, there is significant variation between the groups in the distribution of endemic species across Indochina’s landscape.
Among amphibians, the occurrence of endemic species is correlated with upland and highland forested regions. Within the group, lineages with restricted breeding requirements contain a higher proportion of restricted range or endemic species. Of the 34 Indochinese species in the ranid genera Amolops and Odorrana , both of which require cold, fastflowing water for egg deposition, 59 % are endemic to the region. Only one species from this group, O. chloronota , has a Pan-Asia distribution; however, it appears that this species is, in fact, a complex of multiple species with possible smaller ranges (Stuart et al., 2006b). Similarly, Indochinese species in the rhacophorid genera Kurixalus and Rhacophorus , both of which require vegetation overhanging slow or still water within forests, have an elevated number of endemics (52 %). Two nonexclusive hypotheses likely explain the correlation between restrictive breeding requirements and endemism. First, restricted breeding requirements limit the number of microhabitats a species can exploit. In a region such as Indochina, with its wide variety of elevations and habitats, this will inherently decrease geographic range as compared to species with more permissive breeding requirements. Second, restricted breeding requirements limit effective dispersal opportunities. This increases the likelihood for lineages to become isolated and subsequently diverge, events that may subsequently produce new, possibly restricted-range species.
In contrast, those amphibian families whose members breed in open, slow, or stagnant waters, including temporary sources such as ditches, tire tracks, or the wet rice paddies that are ubiquitous through the region, have the lowest rates of amphibian endemism in Indochina: Bufonidae (0 %), Dicroglossidae (21 %), and Microhylidae (27 %). These lineages also have high numbers of Pan-Asia species compared to the rest of the amphibian fauna: Bufonidae (50 %), Dicroglossidae (28 %), and Microhylidae (36 %). The correlation between slow and/or still water open-habitat breeding requirements and broad geographic distributions is also observed for individual species within other lineages (e.g., Hylarana erythreae , H. taipehensis , Kurixalus verrucosus , Polypedates leucomystax , and P. megacephalus ). For these families and species, the ability to occupy widespread lowland, open habitats increases potential geographic range and reduces the likelihood of isolation and subsequent lineage divergence. If these species are truly widespread and panmictic, then genetic analyses should reveal high levels of gene flow within species across their wide range.
Like amphibians, endemic snakes are significantly more likely to be found in forested environments and within a single altitude zone than their nonendemic counterparts. Unlike amphibians, there is no bias towards an upland distribution; they are more likely to be restricted to either upland or lowland environments. The relationship between microhabitat use and the occurrence of endemic snake species is less clear. Among snakes restricted to a single microhabitat, there is a significant difference between endemics and nonendemics in the type of microhabitat occupied (Fisher’s exact test, p, 0.05; 19 % of endemics and 15 % of nonendemics use multiple microhabitats). The major differences are seen in three groups: more endemics are fossorial (19 % of the endemic fauna vs. 7 % of the nonendemic one) and fewer are terrestrial (34 % vs. 43 %) or arboreal (3 % vs. 19 %) than nonendemic snakes. These results are consistent with recent work in Sundaland suggesting that, at the time scale of Pleistocene climatic fluctuations, terrestrial and arboreal snake species have higher dispersal potential than fossorial ones (Inger and Voris, 2001). In Indochina, snake lineages with the highest rates of endemism in the region are the largely fossorial families Xenodermatidae and Typhlopidae and subfamily Calamarinae . In the Viperidae , which also has a high number of endemic species, three endemics are fossorial ( Protobothrops sieversorum , Protobothrops trungkhanhensis , Viridovipera truongsonensis ) and the remaining one is rupiculous ( Cryptelytrops honsonensis ). Fossorial behavior limits mobility and reduces dispersal opportunities, as individuals may have to go above ground to move substantial distances ( Goodenough et al., 2009). The negative relationship between arboreality and endemism is less clearly related to lineagelevel ecological constraints. Only one strictly arboreal snake, Boiga bourreti , is endemic to Indochina, and the 11 strictly terrestrial endemics are distributed across seven families or colubrid subfamilies.
Endemism among lizards is similar to that of snakes: it is correlated with forested environments at all elevations with individual species generally occupying only a single altitude zone. However, endemic lizards are more strongly associated with lowland environments than are amphibians and the other reptile groups: a relatively large proportion (37 %) are found only in the lowlands and an equal number of endemic species can be found above and below 300 m. As with snakes, fossorial behavior is correlated with restricted range with high rates of endemism observed among the limbless, burrowing family Dibamidae . Endemic lizards are also significantly more restricted than nonendemics to a single microhabitat and the number and type of microhabitats used by endemic species vary with both elevation and lineage. Lowland-restricted endemic lizards are significantly more likely to occupy a single microhabitat than the rest of Indochina’s endemic lizards (x 2 5 10.325, df 5 1, p, 0.005). This suite of lowland endemics is dominated by 17 strictly rupicolous species (61 % of all lowland endemic lizard fauna) all of which are gekkonids with the single exception of the scincid Leptoseps tetradactylus . The high number of lowland endemics wthin this group is likely related to their habitat use. Recent intensive surveys of Indochina’s granitic and limestone karst forests have yielded large numbers of previously undescribed gekkonids of the genera Cnemaspis and Cyrtodactylus occupying remarkably small geographic ranges (e.g., Grismer, 2010; Grismer et al., 2010 b; Ziegler et al., 2010). Many of these species have been discovered in the low-lying South- ern Islands and the low rocky outcrops of the lower Mekong watershed (which act as virtual islands). Although this pattern of highlylocalized, rupiculous endemism has only been recently been recognized, the large numbers of these species occupying small ranges suggest that increased survey effort will have little effect on the reported distributions. The high occurrence of endemism among this group may be related to the isolating nature of such habitats.
Among Indochina’s herpetofauna, traits associated with greater dispersal potential are correlated with reduced endemism and larger geographic ranges and vice versa. In addition to the lineage-specific ecological and behavioral attributes discussed above, it is worth noting a few other factors that may underlie the distribution of Indochina’s endemic species. In general, body size is positively correlated with dispersal distance for actively dispersing organisms (Sutherland et al., 2000; Jenkins et al., 2007). This may contribute to the observation of lower overall endemism rates among snakes, turtles, and crocodiles compared to the smaller, inherently less vagile amphibians and lizards. Additionally, the ability to exploit interconnected and widespread habitats within a given region can also increase an organism’s inherent capacity for movement. In Indochina, reduced rates of endemism are observed among a suite of larger-bodied aquatic or semiaquatic lineages adapted for rapid movement within the region’s widespread network of lowland waterways and wetlands: varanid lizards, freshwater turtles ( Geoemydidae , Tryonichidae), the agamid lizard Physignathus cocincinus , pythons, the aquatic homalopsine and natricine snakes, and both crocodile species.
NONENDEMIC FAUNA: Indochina’s major nonendemic faunal components ( South China, Mainland Southeast Asia , Pan-Asia ) occupy wider ecological niches than the endemic fauna. However , there are significant differences in ecological characteristics among these faunal components that correlate with the geographic ranges they occupy outside Indochina. Regions of South China adjacent to northern Indochina are dominated by forested uplands and highlands and of the South China species known from Indochina, 72 % are forest-restricted and 25 % are found only at elevations above 800 m. Mainland Southeast Asia species restricted to a single elevation are equally likely to be found above and below 300 m (24 %), mirroring the combination of upland and lowland habitats found in adjacent regions of Thailand and Myanmar. Although the distribution of these two faunas (as well as the poorly represented strictly Malaya fauna) across Indochina is largely correlated with geographic proximity, ecological characteristics also contribute to their Indochinese distributions, e.g., South China species in the Central Annamites ; Malaya species in the central and Southern Annamites .
The Indochinese fauna found outside these two regions are dominated by species capable of widespread dispersal: for example, the majority of species in the Pan-Asia fauna tolerate open environments (59 %) and are found at all elevations (60 %). These characteristics allow species to move across relatively large regions, which historically would have included the river-crossed lowlands of the Sunda Shelf exposed during periods of lower sea level as well as higher elevations leading to east Asia and the foothills of the Himalayas.
CHARACTERIZATION OF INDOCHINA’ S HERPETOFAUNA
Our observations are in general agreement with previous research suggesting that Indochina’s floral and faunal diversity is driven in part by the intersection of a northerly subtropical and temperate biota with a southerly tropical one (e.g., Smith, 1931; Bourret, 1936a, b, 1941a, 1942; Bobrov, 1997; MacKinnon, 1997; Averyanov et al., 2003; Orlov, 2005; Sterling et al., 2006). There is a larger presence of species restricted to areas north of Mainland Southeast Asia (South China, Southern Himalayas, East Asia) to Indochina’s herpetofauna is significantly stronger than there is of those restricted to regions to the south (Malaya).
Although there is strong latitudinal variation in the distribution of these two subtropical/temperate and tropical faunas, the transition between the two more closely resembles a threshold than a gradient, with little evidence of true mixing at intermediate latitudes, regardless of elevation. At a finer scale, there is evidence that the Central Annamites harbors small, roughly equivalent numbers of highland-restricted species from the Northeast and/or Northwest Uplands and southern Indochina’s combined upland regions (Southern Annamites, Southern Lao Uplands, Cardamom Uplands). In addition, the substantial endemic Indochinese herpetofauna cannot be easily characterized as temperate, subtropical, or tropical given the region’s complex topography and climate.
Indochina’s geographic location south of China and along the eastern coastal edge of Mainland Southeast Asia likely drives the distribution of nonendemic faunas within the region. The majority of South China species are concentrated in the northern Indochinese uplands adjacent to floristically and climatically similar habitats in China and extreme northern Myanmar, which provide corridors for movement (Nguyen and Harder, 1996; Zhu, 1997; Zhu et al., 2003). Mainland Southeast Asia species are concentrated along Indochina’s western edge. These occur in lower numbers than South China species despite the fact that Indochina shares a longer border with Thailand and Myanmar (2812 km) than with China (1704 km). This may be due to lower species richness and lower dispersal rates across Thailand’s Khorat Plateau, a largely dry, homogeneous plain stretching 900 km along Indochina’s western border. Recent surveys that have filled in distributional gaps for two Pan-Asia lineages, the lizard genus Cnemaspis and pitviper genus Cryptelytrops , suggest that the lower number of Mainland Southeast Asia species in Indochina may also be an artifact of survey effort along Indochina’s southernmost limits ( Grismer and Ngo, 2007; Greismer et al., 2008c).
Historically, the repeated cycles of connectivity and isolation between Mainland Southeast Asia and the Sunda Islands have affected the distribution of Indochina’s nonendemic fauna. Beginning in the mid-Miocene through the present, periods of lower sea level allowed movement across the exposed continental shelf, a lowland region covered with a mosaic of rivers, gallery and evergreen forests, and drier vegetation types. Dispersal across these land bridges is consistent with the observation that the vast majority of Indochina’s lowlandrestricted Pan-Asia species (93 %) also occur in the Sunda Islands. This ecologically tolerant group would have been able to move through the exposed continental shelf fairly easily. The hypothesis that Pan-Asia species have moved frequently and easily between mainland and insular Southeast Asia can be tested by examining the extent of genetic divergence between lineages from the two regions.
The existence of periods of connectivity between Mainland Southeast Asia and the Sunda Islands is also consistent with the presence of apparently relict upland Sundaic species in the Annamites. Of the five strictly Malaya species known from Indochina, three are restricted to forested uplands and known from single locations in the Central Annamites ( Sibynophis melanocephalus , Calamaria lovii ) and Southern Annamites ( Boiga jaspidea ). If the Indochinese populations of these three species are actually distinct species from the Sunda populations, then a sister relationship between island and mainland species would support the hypothesis of allopatric speciation via dispersal across the exposed continental shelf and subsequent isolation during periods of higher sea level. If they are not sister species, further examination of genetic divergence between lineages could be used to infer the dispersal patterns of the Indochinese species.
Within Indochina, the distribution of a subset of the herpetofauna has been positively affected by people. Much of Indochina, particularly the deltas and other lowland subregions, have been vastly altered by human activities for millennia and such activities have contributed to the wide distribution of amphibian and reptile species whose broad ecological tolerances enable them to colonize and persist in human-modified habitats. This group includes species with feeding requirements that are favored in human-dominated environments (e.g., Gekko gecko , Hemidactylus frenatus , Chrysopelea ornata , Coelognathus radiata , Orthriophis taeniura , Ptyas korros , Rhabdophis spp. , Rhamphotyphlops braminus ); species that are insensitive to perturbation of forested areas (e.g., species of Calotes , Lygosoma , Mabuya , Scincella , as well as Theloderma asperum , Bungarus fasciatus , Naja kaouthia , Ophiophagus hannah , Python reticulatus ); and species that are native to open grass and scrublands (e.g., Takydromus sexlineatus , Ahaetulla prasina , Xenopeltis unicolor ). These species are currently widespread across Indochina and elsewhere in Asia; it is unclear if their ranges were more restricted before human colonization. Estimates of genetic diversification between populations could be used to detect the recent rapid population increases expected if their geographic ranges expanded rapidly following human disturbance (Knowles, 2009).
DIRECTIONS FOR FURTHER RESEARCH
Future research will continue to refine and elaborate our understanding of Indochina’s herpetofaunal biogeography and its origins in the region’s complex history. Results from recent efforts clearly indicate that Indochina’s amphibian and reptile richness remains imperfectly known. Between 1997 and August of 2010, increased survey effort and closer examination of natural history collections resulted in the description of 65 new species of amphibians and 77 new species of reptiles from Indochina, as well as new records for 40 amphibian and 36 reptile species previously not known from Indochina. This productivity is consistent with a recent analysis indicating high levels of unknown biodiversity, and suggests that survey efforts are still inadequate (Stuart and Bain, 2008; Giam et al., 2010). Combining current knowledge with trends in survey work and taxonomic and systematic research allows us to postulate what some of the effects of an increased effort would be.
Surveys of greater Southeast Asia, combined with taxonomic work (see below) will likely increase the known relative contributions of the adjacent South China and Mainland Southeast Asia faunas to Indochina’s fauna. It is unlikely that large numbers of widespread species have yet to be identified from Indochina. However, our knowledge of their true ranges within the region will benefit greatly from future surveying; this is particularly true for widespread species whose conservation is of concern, including varanids, crocodilians, pythonids, and freshwater turtles.
Within this context, our analyses of survey efforts indicate that some regions are particularly poorly known: the islands and coasts, as well as the western portions of Indochina, including the Cardamoms, Southern Lao Uplands, and Upper Mekong Lowlands. Efforts in the Cardamoms to date have been weak (15 total visits) and surveys continue to increase their known diversity. The productivity of this work is potentially quite high, for example, the discovery of the last remaining population of Crocodylus siamensis likely reflects a relatively intact herpetofaunal assemblage in the Cardamoms (e.g., Platt and Ngo, 2000; Stuart and Platt, 2000). In addition to increasing estimates of the subregion’s species richness and endemism, future work will clarify the relationship between the Cardamom fauna and that of adjacent Thailand.
Both the flora and fauna of the Southern Lao Uplands, including the Boloven Plateau, remain poorly known. The region has been rigorously surveyed for herpetofauna seven times and remains poorly surveyed for plants (Vidal, 1956–1960), mammals ( Delacour, 1940), birds ( Delacour, 1942, Duckworth et al., 1999), fish (Roberts, 1997, 1998; Kottelat, 2000), and invertebrates ( Gressitt, 1970). This overall lack of knowledge makes it difficult to predict the nature of the subregion’s herpetofauna. We anticipate that current species richness for this upland area is greatly underestimated, especially given its complex topography and high annual rainfall. Increased survey efforts will also define the relationship of the Southern Lao Uplands fauna to the Annamites and to the northern uplands.
Although Indochina’s Northern and Southern Islands have been visited more than average per km 2 when compared to the rest of the region, their insular nature suggests that further surveying is required. Only 13 of the more than 70 islands have been visited to date and multiple surveys have only been conducted on the largest formations: Con Dao, Phu Quoc, and Cat Ba (Smith, 1920; Darevsky, 1999; Grismer and Ngo, 2007; Ziegler et al., 2008c; Grismer et al., 2008c, 2010b; Ngo et al., 2010). Recent descriptions of a gekko in the Northern Islands, as well as gekkonids and a viperid in Rach Gia Bay off southern Vietnam suggest that these areas are not only under- surveyed, but potentially harbor endemic species ( Grismer and Ngo, 2007; Ziegler et al., 2008c; Grismer et al., 2008c, 2010b; Ngo et al., 2010). Given the lack of permanent and diverse freshwater sources, it is unlikely that future surveys will record many more amphibian or freshwater turtle species from these areas.
Relative survey effort could not be estimated for Indochina’s coastal regions; however, it is certain that these areas have been surveyed infrequently (table 2), particularly in the north, and only a few locations have been visited more than once (Smith, 1920; Ziegler and Wetkus, 1999a, b; Das, 2004). There is evidence from both cycads and Dipterocarpaceae that the south-central and southern coast of Vietnam may be areas of elevated endemism associated with extreme aridity (Nguyen and Phan, 1999; Rundel, 2000). A small suite of endemic reptiles restricted to the coasts is known, so increased efforts may further add to the overall number of reptile species, as well as endemic species.
Survey effort is unevenly distributed within subregions as well as among them. To date, 51 out of the 65 surveys conducted in the Northwest Uplands have been restricted to Vietnam, which makes up less than half of this subregion and has much smaller borders with regions outside Indochina than does northern Laos. As more surveys are undertaken in northern Laos, we expect recorded species richness to increase, including endemic species and those currently known from southwestern Yunnan, northern Thailand, and northern Myanmar. For some subregions, only a few localities account for a large proportion of the overall survey effort: Nha Trang, 37 % of all South Coast surveys; Da Nang, 58 % of all Central Coast surveys; Ho Chi Minh City, 25 % of all Mekong Delta surveys; Vientiane, 36 % of all Upper Mekong lowlands surveys; Tam Dao, 45 % of all Northeast Uplands surveys; and Fan Si Pan, 37 % of all Northwest Uplands surveys (appendix 2A). More locality-balanced survey work will have a great impact on determining species richness within these subregions in particular.
Future survey work will also refine Indochina’s biogeographic subunits as defined by its herpetofauna and answer questions that currently cannot be resolved. The strong amphibian and reptile faunal overlap between South China and the upland subregions of northern Indochina suggests that future research, and especially increased international collaborations, will help to resolve the borders of this region as a single biogeographic unit. Strong current similarities between the Cardamoms and Mainland Southeast Asia faunas suggest that additional surveys are needed to clarify how closely this region is allied with the rest of Mainland Southeast Asia. Vietnam’s northeast and Central–South Vietnam Lowlands may contain more substructure than our subregional classification was able to capture, given the limited data available. Similarly, the Annamites may have more substructuring than we outline in our a priori boundaries of its three focal subregions. In particular, the faunal relationship between the Annamites and the Southern Lao Uplands remains to be clarified. Finally, characterizing the relationships between the Central–South Vietnam Lowlands and its associated uplands (Central and Southern Annamites) and lowlands (low- er Mekong watershed) may help us to understand the factors affecting the distribution of endemic species in southern Indochina. More detailed distribution data will also allow us to map distributions for a sufficient number of species to test biogeographic hypotheses and refine the boundaries of our largely a priori subregions.
Future taxonomic work is also greatly needed in the region. Cross-border taxonomic issues (primarily with China) clearly bias our estimates of species richness and endemic species numbers. There are 78 species of amphibians and reptiles whose distributions terminate at the international border between Indochina and South China: 38 species distributed through China up to the Vietnam and/or Lao border, but not within Indochina (table 12) and 40 species that are distributed through Indochina up to the Chinese border, but not within China (table 13) (after Daming, 2002; Zhou and Yu, 2002; Ye et al., 2007; Fei et al., 2008, 2009; see appendix 2A). These ranges are the result of the parallel systematic work historically undertaken on each side of the border, with little communication or cooperation. This is significant because these international borders are not concomitant with any geographic feature and represent a taxonomic artifact that biases biogeographic analyses. It is likely that a number of these species belong to the large group restricted to South China and northern Indochina; only nine species are known from outside these regions, eight from Mainland Southeast Asia and one from Pan-Asia, a pattern that mirrors the large faunal sharing between northern Indochina and South China. Furthermore, species not yet recorded from border areas have also been shown to exist across these international borders (e.g., Bain and Stuart, 2006a), suggesting that the extent of the cross-border taxonomic problem as presented above is an underestimation. Increased cross-border communication, will result in a decrease of the known regional richness, concomitant with an increase in synonymy, and a decrease in known Indochinese endemism.
At the same time, increased taxonomic and systematic work will likely increase estimates of Indochina’s species richness and endemism. Certain taxonomic groups are underrepresented in the region’s current species list. These include fossorial species that often evade detection in surveys, such as ichthyophiids, anguids, dibamids, and calamarines; strictly arboreal species that can also evade detection; species that are part of poorly defined taxonomic groups such as those of the genera Dinodon (see Orlov and Ryabov, 2004), Oligodon (see David et al., 2008b, c), Amphiesma (see David et al., 2007), and Draco (see McGuire and Heang, 2001). Combined survey efforts and morphological and molecular analyses will likely resolve some of the wide-ranging cryptic species complexes such as those of the genera Paramesotriton (see Chan et al., 2001), Fejervarya (see Toda et al., 1998a, b; Veith et al., 2001), Limnonectes (see Emerson et al., 2000; Evans et al., 2003; McLeod, 2010), Odorrana (see Fei et al., 2001; Li et al., 2001; Bain et al., 2003, 2009c; Orlov et al., 2006c), Hylarana (see Murphy et al., 1997; Matsui et al., 2002), Polypedates (see He, 1999; Orlov et al., 2002b; Bain and Nguyen, 2004b), Rhacophorus (see Ohler and Delorme, 2006), Calotes (see Zug et al., 2006), Leiolepis (see Schmitz et al., 2001), Takydromus (see Lin et al., 2002), Dixonius (see Ota
TABLE 12 Herpetofauna of China with ranges ending at the northern border of Indochina See Materials and Methods for definition of ranges.
et al., 2001), Emoia (see Emilio et al., 1996), Naja (see Wüster and Thorpe, 1992), possibly Sacalia (Shi et al., 2008) , and several genera of green tree vipers (see Malhotra and Thorpe, 2004a, b). This will result in an increase in the number of endemic species and a concomitant decrease in the number of widespread species. Resolution of these complexes will also provide phylogenetically based insights into ecological, geographic, and historical barriers structuring regional diversity.
TABLE 13 Herpetofauna of Indochina with ranges ending at the northern border with China See Materials and Methods for definition of ranges.
CONCLUSIONS
Our analysis provides the first modern synthesis of Indochina’s biogeography. Research to date clearly indicates that amphib- ian and reptile species of Indochina represent a globally significant fauna, given its high levels of endemism, significant presence of species found outside Mainland Southeast Asia, and strong differentiation among internal subregions. The composition of this herpetofauna reflects an intricate evolutionary history that has been strongly influenced by the region’s complex geological and climatic history. Despite these high levels of species richness and endemism, Indochina is still insufficiently surveyed.
We have made some preliminary assessments of the structure of regional diversity. We have also outlined areas in need of refinement and further testing, including: (1) assessments of the degree of substructure in the Annamites, lowlands of centralsouth Vietnam, and the lower Mekong watershed; (2) the relationship of the Southern Lao Uplands herpetofauna to the rest of Indochina’s uplands; (3) the divergence of disjunct Indochina-Sunda populations; (4) the presence and structure of a restricted-range northern Indochina/ South China herpetofauna; and (5) clarification of the biogeographic barrier separating northeastern Vietnam from northwestern Vietnam and northern Laos. Finer resolution mapping (resulting in part from increased survey efforts), alpha-level taxonomic work, and phylogenetic analyses at a regional level will assist in elucidating both distributional patterns and the evolutionary histories contributing to current patterns of diversity.
The unique character of the region and its herpetofauna makes it a vital focus for conservation efforts, given the current intensity of threats to Indochina’s natural resources. Currently, 41 amphibian and 23 reptile species included in this review are classified as globally threatened, of which 29 (17 amphibians, 12 reptiles) are endemic to the region of Indochina and South China (IUCN, 2010). This survey provides data on amphibian and reptile species richness and distribution that are necessary for the development of regional and global conservation priorities. Effective conservation efforts require that this material be integrated with data on threat levels to the herpetofauna; consequently, the development of a database of threatened amphibian and reptile species is now a major priority in the region. Synthesizing data on conservation status with the above biogeographic analyses will help elucidate the patterns of threat to the Indochinese herpetofauna and enable this fauna to be incorporated into regional conservation planning.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.